Механизм образования трансдуцирующих фагов, осуществляющих специфическую трансдукцию
Специфическую трансдукцию осуществляют фаги, ДНК которых при лизогенном пути развития включается в бактериальную хромосому в специфическом сайте и реплицируется вместе с ней. К таким фагам относятся лямбоидные фаги E. coli. Наиболее хорошо изучен фаг лямбда (λ).
Интеграция ДНК фага лямбда происходит через специфические att-сайты (от англ. аttachment), имеющиеся на фаговой и бактериальной ДНК. Фаговый (attР-РОР') и бактериальный (аttВ-ВОВ') сайты устроены сходным образом. Их основу составляют одинаковые последовательности из 15 нуклеотидов (О), фланкированные участками определенного строения. Эти участки у фагов обозначаются Р и Р', у бактерий – В и В'. Участки Р и Р' состоят соответственно из 145 и 75, а В и В' – из 3 - 5 п.н. Специальный фаговый белок делает ступенчатые разрывы в участке О фаговой и бактериальной ДНК, с образованием однонитевых комплементарных участков из 7 нуклеотидов. При взаимном обмене такими концами между РОР' и ВОВ' фаговая ДНК встраивается в бактериальную хромосому. Причем на концах профага оказываются рекомбинантные att-сайты ВОР' и РОВ' (рис.15). В случае индукции профаг исключается через эти сайты из бактериальной хромосомы.

|
Рис.15. Встраивание фаговой ДНК в бактериальную хромосому.
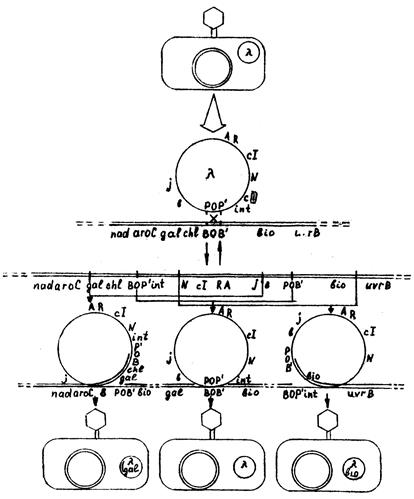
Изредка, с частотой около 10-6, исключение профага осуществляется по случайным сайтам с захватом бактериальных генов, располагающихся слева или справа от него. Эти гены локализуются в фаговой ДНК соответственно слева или справа от attРОР' сайта. Такой фаг будет переносить при инфекции реципиентной клетки только определенные гены донора. Следовательно, фаги, вызывающие специфическую трансдукцию, являются рекомбинантами, содержащими фаговую и бактериальную ДНК. Специфическая трансдукция наблюдается только тогда, когда фаговые лизаты получены при спонтанном или индуцированном освобождении фага из лизогенизированного донора.
|

|
Рис.16. Механизм образования трансдуцирующих фагов, осуществляющих специфическую трансдукцию (на примере фага λ)
Молекулы ДНК, длина которых превышает длину ДНК фага лямбда. (48502 п.н.) более чем на 5-6% не упаковываются в головку. Поэтому включение бактериальных генов в фаговый геном происходит при потере части фаговой ДНК, что часто вызывает дефектность трансдуцирующих фагов. В дефектные трансдуцирующие фаги вместо генов левой части фагового генома может включиться до 25 т.п.н., а с другого конца только до 12 т.п.н., так как в нем локализуются гены, осуществляющие автономную репликацию ДНК фага. Если произойдет замена генов, несущественных для вегетативного развития фагов (не более 10 т.п.н.) и группирующихся вместе на обоих концах профага, то образуются жизнеспособные трансдуцирующие фаги.
Обычно фаг лямбда может трансдуцировать гены, удаленные от attВ сайта не более чем на 10-20 т.п.н., но с помощью разных молекулярно-генетических методов было выделено более 100 трансдуцирующих фагов лямбда, которые несут бактериальные гены, расположенные в разных частях хромосомы.
Судьба перенесенного фрагмента хромосомы различна. Трансду-цированный фрагмент может остаться соединенным с генетическим материалом фага и при встраивании фага в ДНК бактерии (лизогенизации бактерии) трансдуцированный фрагмент будет представлен в мерозиготе двумя копиями (донорной и реципиентной). Такое мерозиготное в отношении трансдуцированного гена состояние (гетерогенота) нестабильно. При размножении мерозигот с частотой 1 х 103 происходит выщепление клеток с генотипом реципиента (за счет потери добавленного фрагмента) или стабильных рекомбинантов (при включении трансдуцированных генов в бактериальную хромосому). Включение фрагмента за счет двойного кроссинговера с образованием стабильных рекомбинантов происходит в 30% случаев.
Дата добавления: 2015-02-07; просмотров: 2269;