Трансформация. Общая характеристика процесса
Трансформация – способ передачи генетической информации от клеток-доноров к клеткам-реципиентам с помощью химически чистой ДНК.
Явление трансформации впервые было обнаружено у пневмококков еще в 1928 году Ф.Гриффитсом. Из его опытов следовало, что погибшие от нагрева (60-80°С) клетки вирулентного пневмококка могли передавать свои вирулентные и антигенные свойства живым авирулентным клеткам другого серотипа. Только в 1944 году 0.Звери, К.МакЛеод, М.МакКарти доказали, что это превращение – трансформацию – осуществляет не белок, не полисахарид, а высокополимерная ДНК вирулентного штамма (как было показано позднее, ДНК сохраняет свою двуспиральную структуру и полимерность при описанных условиях тепловой обработки). В 1957 году Л.Лерман и Л.Толмач непосредственно продемонстрировали, что клетки бактерий способны поглощать нативную (двухцепочечную) полимерную молекулу ДНК.
Трансформация в настоящее время воспроизведена на многих видах бактерий, число которых непрерывно умножается, и является основным методом введения векторной ДНК в клетки бактерий при генно-инженерных работах. Для некоторых видов (пневмококков, стрептококков, сенной палочки и гемофильных бактерий) она является естественным способом обмена генетической информацией, выработанным в процессе эволюции. Для других (бактерии кишечной группы) такой способ обмена не происходит в естественных условиях, но его можно воспроизвести в лаборатории. Трансформацию удалось получить не только между бактериями одного вида, но также и между бактериями, принадлежащими к разным видам.
Наилучшей трансформирующей активностью обладает нативная (двуспиральная) и высокополимерная ДНК, молекулярная масса которой от 10 и более мегадальтон (МДа). Величина участка ДНК, еще обладающего трансформирующей активностью, составляет 0,3 МДа. Верхний предел фрагмента, который может поглотить клетка, очень высок. ДНК с молекулярной массой 500 МДа из гемофильной бактерии (половина всей хромосомы) обладала даже большей трансформирующей активностью, чем обычные препараты в 15-20 МДа. Трансформирующая активность денатурированной (одноцепочечной) ДНК составляет 0,5 ~ 2,0% от активности нативной ДНК.
В процессе трансформации у всех видов бактерий можно условно выделить четыре этапа: 1) адсорбция и поглощение клеткой молекулы ДНК; 2) эклипс-период; 3) интеграция поглощенной ДНК в геном реципиента; 4) фенотипическое выражение признаков, детерминированных перенесенным фрагментом.
Механизм поглощения молекул ДНК различается у разных групп бактерий, т.е. он видоспецифичен. Трансформация наиболее хорошо изучена у пневмококков, близких к ним стрептококков, сенной палочки и гемофильных бактерий.
Все клетки популяции способны адсорбировать ДНК на клеточной стенке, но не все способны ее поглощать. Бактериальные клетки, способные поглощать ДНК, называются компетентными. Культуру, содержащую такие клетки, называют культурой в стадии компетентности. У большинства бактерий компетентность возникает лишь на определенном этапе роста. Культура стрептококков в этой стадии находится на этапе логарифмического роста, культуры других бактерий – на более поздних этапах. У гонококков и менингококков культура компетентна в любой стадии роста. У E. coli естественная компетентность вообще отсутствует, и поглощение ДНК происходит лишь при определенной обработке клеток.
В стадии компетентности у пневмококков, стрептококков, гонококков и гемофильных бактерий компетентна практически каждая клетка в культуре. У сенной палочки даже в период максимальной компетентности к поглощению ДНК способны лишь 10 - 15% клеток популяции.
Компетентные клетки отличаются от некомпетентных не только способностью поглощать ДНК, но и другими свойствами. Они обладают сниженным уровнем метаболизма и темпом репликации ДНК. Изменены свойства наружных слоев клетки, снижен заряд клеточной поверхности. Они более чувствительны к осмотическому шоку, тепловой обработке. Устойчивее к пенициллину.
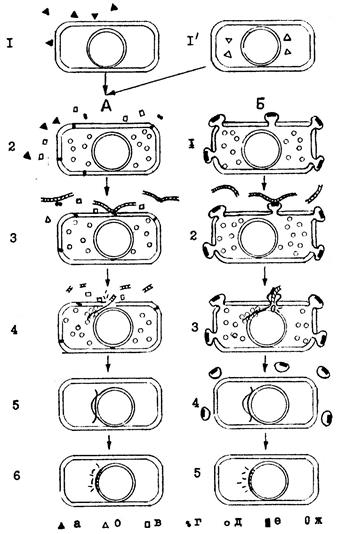
Особенности развития компетентности, этапы поглощения трансформирующей ДНК клетками пневмококков, стрептококков, сенной палочки (А) и клетками гемофильных бактерий (Б) схематично изображены на рис.11.
|
Рис.11. Этапы поглощения трансформирующей ДНК клетками (А) пневмококков, стрептококков, сенной палочки и (Б) гемофильных бактерий (объяснение в тексте).
А: 1 – пневмококки, стрептококки. Часть клеток в культуре начинает синтезировать и выделять в среду фактор компетентности, или активатор (а), он адсорбируется на остальных клетках и индуцирует у них компетентность. У пневмококков достаточно 0,1 мкг на мл фактора для превращения 107 клеток в компетентные за 15-20 минут. 1'- сенная палочка. У части клеток в культуре происходит смена синтеза одной РНК-полимеразы на другую (б). Это изменение, ведущее к возникновению у клетки состояния компетентности, охватывает не более 15-20% всех клеток данной культуры. У сенной палочки компетентность связана с нарушением целостности клеточной стенки. Часть клеток в популяции превращается под действием автолитических ферментов или факторов, способствующих к несбалансированному росту бактерий, в частичные протопласты.
2 – фактор компетентности, после адсорбции на клетке стрептококков и пневмококов, или появление нового типа РНК-полимеразы в клетках сенной палочки "включает" синтез ряда новых белков. К ним относятся аутолитические ферменты (в), белковый комплекс, способный связываться с трансформирующей ДНК и приводить к однонитевым и двунитевым ее разрывам (г), белки, участвующие в рекомбинации (д), сам фактор компетентности (а). Некоторые из этих белков находятся в периплазматическом пространстве и после частичного разрушения клеточной стенки выходят во внешнюю среду.
3 – трансформирующая ДНК связывается соответствующим белковым комплексом (г) и адсорбируется на поверхности клеток. Необратимая адсорбция ДНК на компетентных клетках происходит в течение нескольких секунд после ее добавления к компетентным клеткам. Фрагмент вначале прикрепляется к клетке несколькими точками. На клеточной стенке пневмо- и стрептококков имеется от 30 до 80 рецепторов прикрепления ДНК, а у сенной палочки – от 20 до 50. Уже в процессе адсорбции примерно через 30 сек высокомолекулярная ДНК расщепляется нуклеазами на двунитевые фрагменты, имеющие молекулярную массу 1-6 МДа (1,5-8 т.п.н.). Эти фрагменты входят в клетку с одного конца. Скорость вхождения примерно 55 нуклеотидов в секунду. Одна клетка пневмококков поглощает в 2-10 раз больше ДНК, чем ее содержится в геноме. У сенной палочки поглощаться может 3-6 молекул с молекулярной массой 16 МДа каждая. Видовая принадлежность ДНК не влияет на ее поглощение клетками пневмококков, стрептококков.
Б (гемофильные бактерии): 1 – в стадии компетентности на поверхности всех клеток культуры появляются выпячивания цитоплазматической мембраны – трансформосомы, диаметром около 80-100 мкм, в количестве 5-13 на клетку. На трансформосомах содержится белок (е), способный «опознавать» специфические последовательности нуклеотидов на ДНК и связываться с ними. Клетки становятся компетентными.
1 – фрагменты ДНК, имеющие специфическую последовательность в 11 нуклеотидов (AAGTGСGGТСА), адсорбируются и необратимо связываются при участии белка на трансформосомах. Таких последовательностей на хромосоме гемофильных бактерий – 600, поэтому лучше связывается гомологичная или родственная ДНК.
2 – ДНК, адсорбированная на трансформосомах, за 3-5 сек втягивается в клетку через пору в цитоплазматической мембране. Каждая трансформосома поглощает лишь одну молекулу ДНК. Общее количество ДНК, поглощаемое компетентной клеткой, равно 5 фрагментам с молекулярной массой 10 МДа каждый. После поглощения трансформосомы отваливаются и стадия компетентности заканчивается.
Эклипс-период или эклипс-фаза – это стадия, предшествующая собственно включению ДНК в хромосому. У всех изученных бактерий процессы превращения трансформирующей ДНК в клетке-реципиенте имеют много общего. Комплекс ДНК-белок проникает сквозь цитоплазматическую мембрану (или пору) внутрь клетки. При этом ДНК становится однонитевой за счет разрушения нуклеазами (ж) второй нити. Обе комплементарные нити ДНК генетически активны и избирательного разрушения одной из них не происходит. С такой однонитевой ДНК связываются белки, участвующие в рекомбинации (д) и одновременно играющие роль протекторов. Система рестрикции-модификации клетки-реципиента не влияет на трансформирующую активность чужой ДНК, так как она либо быстро вступает в синапсис и рекомбинирует с ДНК хромосомы реципиента, либо не доступна из-за однонитевого состояния. Эклипс-фаза у сенной палочки длится при 30°С 20 минут, при 37°С – 10 минут.
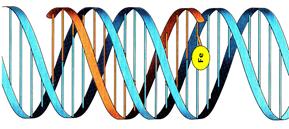
Линейная трансформирующая ДНК не может существовать и функционировать в клетке на протяжении нескольких поколений в автономном виде. На следующем этапе – этапе интеграции – происходит включение однонитевого фрагмента трансформирующей ДНК в гомологичную область хромосомы реципиента, с помощью процесса, получившего название – конверсия. Конверсия гена - нереципрокная гомологичная рекомбинация, основанная на коррекции неспаренных оснований в рекомбинационном гетеродуплексе. Вначале однонитевой трансформирующий фрагмент ДНК соединяется за счет водородных связей с одной из нитей дуплекса реципиента, вытесняя вторую нить. Формируется трехнитевой участок – так называемая, Д-петля(рис.12).
|
Рис.12. Формирование Д-петли.
Затем замещенный участок ДНК реципиента выбрасывается из хромосомы и разрушается до нуклеотидов, а встроенная нить донорной ДНК закрепляется ковалентными связями. В этом процессе участвуют белки рекомбинации. В результате формируется гетеродуплексный участок, в котором одна нить в молекуле ДНК от донора, а вторая от реципиента. Если в этом участке отсутствует полное соответствие азотистых оснований, то он подвергается коррекции. Она заключается в выщеплении одной из нитей, принадлежащих донору или реципиенту, из двунитевого участка ДНК и репаративном синтезе новой нити, полностью комплементарной оставшейся на месте образовавшейся бреши нити-матрице.
Включение двух комплементарных нитей донора определяет случай: чаще это происходит лишь с одной нитью, но может быть одновременное включение обеих.
Фенотипическое выражение признака происходит лишь после включения бактериальной трансформирующей ДНК в хромосому реципиента. Время появления признака зависит от вида бактерий и от перенесенного маркера. У пневмококков фермент, расщепляющий мальтозу, появлялся в реципиентной клетке через 10 минут после поглощения ДНК донора, содержавшей нормальную аллель этого гена, а фермент гистидаза – через 45 минут. Для проявления устойчивости к антибиотикам необходимо 60-120 минут. Подвижность клеток сенной палочки проявлялась через три часа после поглощения ДНК.
Дата добавления: 2015-02-07; просмотров: 3228;