Рабочий цикл актомиозинового комплекса
В процессе сокращения мышцы каждая головка миозина совершает многократные повороты, периодически изменяя угол своего наклона относительно нити актина. В расслабленной мышце миозиновый мостик отделен от актиновой цепи (рис. 4, состояния 1 и 2). Свободная головка миозина обладает определенной степенью подвижности; за счет шарниров, расположенных в местах соединения фрагментов S1 и S2, угол ее наклона относительно хвоста может изменяться. При связывании молекулы АТР с активным центром миозина его головка остается отсоединенной от актина (рис. 4, состояние 1). В каталитическом центре миозина молекула АТР расщепляется на ADP и Pi (рис. 4, переход "состояние 1" à "состояние 2"). Образующиеся при этом молекулы ADP и Pi остаются прочно связанными с каталитическим центром. Однако вслед за гидролизом молекулы АТР происходит присоединение головки миозина к актиновой нити: сначала образуется слабая связь (состояние 3), затем возникает более прочная связь (состояние 4). При этом вращательная подвижность миозинового мостика становится ограниченной. Прочное связывание головки миозина с актином инициирует освобождение фосфата Рi из активного центра (переход "состояние 4 " à"состояние 5 "). Изменения, происходящие в каталитическом центре после диссоциации Рi , вызывают дополнительное увеличение сродства миозина к актину. В результате этого появляется сила, вызывающая поворот мостика в сторону хвоста (переход "состояние 5 "à "состояние 6 "). Вместе с поворотом мостика смещается вдоль нити актина хвост миозина, который соединен с мостиком с помощью "шарнирного" сочленения. Благодаря продольному смещению хвоста миозина происходит сокращение длины саркомера. После смещения головки миозина, инициированного диссоциацией фосфата, молекула ADP диссоциирует из каталитического центра, а ее место занимает новая молекула АТР (переход "состояние 7 "à "состояние 1 "). Это превращение сопровождается отсоединением головки миозина от актина, завершающим цикл структурных преобразований, происходящих в активном центре миозина. В результате многократно повторяющихся циклов гидролиза АТР возникает направленное скольжение нитей миозина и актина друг относительно друга.
Кооперативная и "индивидуальная трудовая деятельность" миозина
В мышечных волокнах молекулы миозина работают не индивидуально, а кооперативно, в составе крупных макромолекулярных ансамблей. Как мы уже отмечали, в миофибриллах молекулы миозина собраны в жгуты, из которых выступает множество миозиновых мостиков в сторону нитей актина (см. рис. 1). Установлено, что во время сокращения мышцы лишь сравнительно небольшая часть мостиков (10-15%) одновременно находится в контакте с окружающими их нитями актина. Это значит, что каждый из мостиков большую часть времени проводит в свободном состоянии, когда он непосредственно не создает тянущей силы. При этом молекулы миозина, у которых мостики отсоединены от актиновых нитей, во время сокращения саркомера перемещаются вместе с остальными молекулами миозинового жгута. Такое движение свободных (несвязанных) мостиков происходит за счет работы других мостиков, которые в это время непосредственно взаимодействуют с нитями актина. Это значит, что каждый мостик не просто шагает вдоль нити, равномерно ступая между соседними звеньями актиновой цепи, а как бы прыгает вдоль нее. Длина таких прыжков составляет 36-38 нм (рис. 5, а), что многократно превышает размер индивидуального шага (Ds ~ 4 нм). Таким образом, сокращение мышечных волокон обеспечивается за счет кооперативной работы большого количества молекул миозина, собранных в толстые нити.
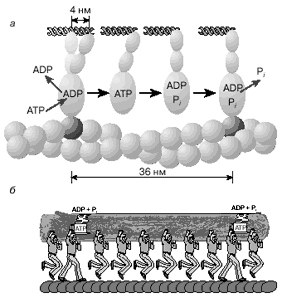
Рис. 5. Схема перемещения молекулы миозина вдоль нити актина.
Движение пучка молекул миозина вдоль нити актина можно сравнить с перетаскиванием бревна большой группой работников, из которых лишь небольшая часть тружеников (10-15%) опирается ногами на землю, в то время как остальные работники в это время висят на бревне, не касаясь земли. Подобно мостикам миозина, работники периодически меняются ролями, однако в каждый момент времени активно работает лишь небольшая часть тружеников, несущих бревно вместе с висящими на нем товарищами (рис. 5, б)
Кооперативный способ работы молекул миозина, характерный для скелетных мышц, встречается также в некоторых других сократительных системах. Известна, однако, большая группа моторных белков, которые работают индивидуально; цикл их механохимических превращений протекает в равномерном режиме, а перемещение происходит отдельными шагами, без длинных прыжков, характерных для миозина скелетных мышц. К таким молекулярным моторам относятся некоторые классы внемышечных миозинов, работающих в клетках животных и растений в качестве перевозчиков мембранных частиц. Простейшие молекулы миозина, выполняющие работу индивидуальных переносчиков (например, миозин класса I), имеют глобулярную головку и короткий хвост. Двигаясь вдоль нити актина цитоскелета, молекула-переносчик может тащить за собой органеллу, к которой она прикрепляется своим хвостом. Таким способом молекулы миозина обеспечивают работу транспортной системы, предназначенной для переноса различных частиц и органелл в клетке.
Движение органелл легко наблюдать в некоторых крупных клетках, таких, как гигантские клетки зеленой водоросли Nitella. Интересна структурная организация транспортной системы, обеспечивающей циркуляцию цитоплазмы в клетках зеленых водорослей. На периферии клетки, рядом с плазматической мембраной, находится слой хлоропластов. Хлоропласты примыкают к так называемому кортикальному слою, который содержит пучки актиновых нитей. Между неподвижными кортикальным слоем и мембраной вакуоли расположен слой движущейся цитоплазмы, в котором находятся ядра, митохондрии и другие органеллы. Молекулы миозина совершают круговое движения внутри клетки, равномерно перемещаясь вдоль нитей актина и увлекая за собой сравнительно крупные органеллы (митохондрии, эндоплазматический ретикулум, ядра и др.). Скорость перемещения органелл составляет 50-75 мкм/с. Благодаря такому движению в гигантской клетке водоросли возникает циркуляция цитоплазмы, за счет которой ее содержимое перемешивается гораздо быстрее, чем это происходило бы путем простой диффузии. Гигантские клетки зеленых водорослей достигают длины 2-5 см, поэтому без принудительной циркуляции цитоплазмы молекулам белков потребовалось бы около 10 дней для диффузии с одного конца клетки на другой.
Дата добавления: 2015-02-07; просмотров: 2418;