АНАТОМИЯ
Нейрогипофиз представляет собой удлиненное выпячивание вентрального гипоталамуса, непосредственно контактирующее с задней и дистальной частью аденогипофиза [I]. У взрослых мужчин и женщин масса нейрогипофиза составляет приблизительно 100 мг, и его можно разделить на две части, соединенные короткой ножкой, проходящей сквозь диафрагму турецкого седла. Верхнюю часть называют по-разному: воронкой или срединным возвышением, а нижнюю — инфундибулярным отростком или pars nervosa. Обе части снабжаются кровью из ветвей верхней и нижней гипофизарных артерий, отходящих от задней коммуникантной и внутрикавернозной части внутренних сонных артерий. В pars nervosa артериолы распадаются на местные капиллярные сети, которые дренируются непосредственно в яремную вену через венозные синусы (турецкого седла, кавернозные и боковые). В воронке первичные капиллярные сети сливаются в другую си-
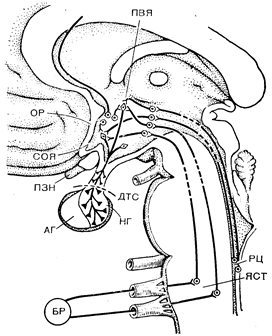
Рис. 9—1. Нейрогипофиз и его основные нервные связи.
НГ — нейрогипофиз; АГ — аденогипофиз: ДТС — диафрагма турецкого седла; ПЗН — перекрест зрительных нервов: СОЯ — супраоптическое ядро: ПВЯ — паравентрикулярное ядро ОР — осморецептор; БР — волюм- и барорецептор ЯСТ — ядро солитарного тракта: РЦ — рвотный центр.
стему — воротные вены, которые перед впадением в системную циркуляцию перфузируют аденогипофиз.
Под микроскопом нейрогипофиз выглядит как густо переплетенная сеть капилляров, питуицитов и безмякотных нервных во-лолон, содержащих большое число электронно-плотных гранул. Эти нейросекреторные нейроны берут начало главным образом в крупноклеточных ядрах супраоптической и паравентрикулярной областей гипоталамуса [2]. Большинство аксонов направляется отсюда вентрально и каудально и оканчивается луковицеобразными расширениями на капиллярных сетях, пронизывающих все части нейрогипофиза, в том числе ножку и воронку (рис. 9—1). Последняя служит местом окончания и других нейросекреторных нейронов, влияющих на переднюю долю гипофиза, путем высвобождения стимулирующих или тормозящих факторов в воротные вены. Играют ли аналогичную роль нейрогипофизарные нейроны воронки — неизвестно. Существует и особый еще меньший отдел нейрогипофиза, который проецируется на III и, вероятно, боковые желудочки мозга [1, З]. По-видимому, из него секрет, выделяется непосредственно в СМЖ, но окончательно его функция не установлена [4].
ХИМИЯ
Единственно известными гормонами, секретируемыми нейрогипофизом взрослого человека, являются вазопрессин и окситоцин. Их структура впервые была расшифрована du Vigneaud с сотр. более 20 лет назад [5]. Каждый из этих гормонов представляет собой нонапептид, состоящий из шестичленного кольца, замкнутого дисульфидным мостиком, и трехчленной концевой части, терминальная карбоксильная группа в которой амидирована (рис. 9—2). Вазопрессин отличается от окситоцина только присутствием фенилаланина на месте изолейцина в кольце и аргинина на месте лейцина в концевой части молекулы. Эти два гормона обнаружены у всех млекопитающих, за исключением подотряда Suina, у некоторых видов которого образуется вариант вазопрессина, содержащий в 8-й позиции лизин вместо аргинина [6]. Окситоцин обнаружен также у многих видов птиц, пресмыкающихся, амфибий и костистых рыб. Однако вместо вазопрессина гипофиз позвоночных, не относящихся к классу млекопитающих, содержит аргининвазотоцин. Его структура отличается от структуры вазопрессина только присутствием в 3-й позиции изолейцина, а биологические эффекты обоих соединений одинаковы. Поскольку вазотоцин — это единственный нонапептидный гормон, находимый у некоторых филюмов древних позвоночных, его считают предшественником, из которого в процессе эволюции вследствие мутации и удвоения генов образовались окситоцин и вазопрессин.
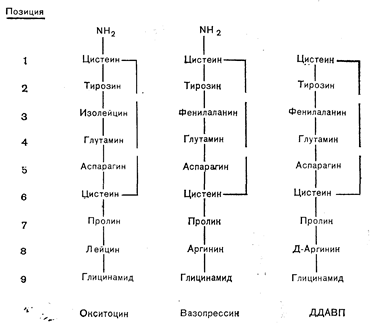
Рис. 9—2. Аминокислотная последовательность окситоцина, вазопрессина и синтетического аналога— 1-дезамино-8-D-аргинин-вазопрессина (ДДАВП).
Синтез большого числа структурных аналогов вазопрессина и окситоцина позволил точнее выяснить зависимость между их конформацией и биологической активностью [7]. Хотя изменение почти любой части молекулы может сказываться на трехмерной структуре и биологической активности, наиболее важными для узнавания и связывания гормона его рецептором считают боковые цепи в 3, 4, 7 и 8-й позиции, тогда как замещения в 5-й позиции влияют на собственную активность молекулы. При замене в 8-й позиции L-аргинина на D-аргинин отношение антидиуретического эффекта вазопрессина к его прессорной активности резко увеличивается. Эта модификация наряду с удалением терминальной аминогруппы цистеина образует 1-дезамино-8-D-аргинин-вазопрессин (ДДАВП) (см. рис. 9—2) — клинически применяемый аналог, обладающий более длительной и сильной антидиуретической активностью [8].
Вазопрессин и окситоцин откладываются в нейрогипофизе в виде нерастворимых комплексов с белками-носителями, получившими название нейрофизинов. Acher и сотр. [9] впервые отделили эти нейрофизины от активных гормонов более 20 лет назад. Однако лишь недавно удалось получить отдельные нейрофизины в достаточно чистом виде, чтобы определить их физикохимические свойства [10]. У человека и большинства млекопитающих иммунологически и хроматографически выделены нейрофизины двух основных типов. Один из них находится исключительно в гранулах, содержащих окситоцин, а второй — только в гранулах, содержащих вазопрессин. Оба представляют собой, по-видимому, одноцепочечные полипептиды с исходной молекулярной массой примерно 10000, но в концентрированных растворах легко образуют димеры и тетрамеры. Нейрофизин каждого типа одинаково хорошо связывает окситоцин и вазопрессин. Это свидетельствует о том, что специфические связи с гормонами, обнаруживаемые in vivo, обусловлены анатомической компартментализацией. Оптимум рН для связывания гормонов с нейрофизином составляет 5, 2—5, 8,. а К—приблизительно 2-105. Такие параметры обеспечивают полную диссоциацию комплекса нейрофизин — гормон в плазме. Частично определена аминокислотная последовательность ряда нейрофизинов, причем выявлена значительная внутренняя гомология той части молекулы, которую считают местом связывания гормонов.
Дата добавления: 2015-01-19; просмотров: 1637;