Цитогенетика и материальные основы наследственности.
Согласно положению клеточной теории (Рудольф Вирхов), новая клетка может появиться только из клетки. Новое поколение при делении клетки от своих родителей получает все признаки, формирующие его организм. Изучением строения материальных структур наследственности и изменчивости и их функционированием занимается особый раздел генетики – цитогенетика.
Основной предмет исследований цитогенетики — хромосомы, их организация, функционирование и наследование. При классическом цитогенетическом анализе проводят одновременно цитологическое (микроскопическое) исследование хромосом и генетический анализ наследования признаков. Цитогенетику подразделяют на общую, в которую включают также популяционную и радиационную цитогенетику, и частную — цитогенетику растений, цитогенетику животных и цитогенетику человека (в том числе медицинскую цитогенетику).
Однако, в передаче наследственных признаков участвуют не только хромосомы, но и другие клеточные структуры. Такое наследование называется внехромосомной (цитоплазматической) наследственностью и ее изучает раздел генетики, называемый клеточной генетикой. При изучении материальных основ наследственности используются не только методы генетики и цитологии, но, и методы молекулярной биологии, цитохимии, кариологии и др.
Роль хромосом в наследовании признаков была определена далеко не сразу, да и установление того факта, что в каждой клетке живого организма обязательно присутствие хромосом заняло продолжительный период. Первые исследования хромосом начались более 100 лет назад с помощью обычного светового микроскопа.
Сейчас сложно сказать, кто сделал первое описание и рисунок хромосом. В 1872г. швейцарский ботаник Карл Вильгельм Нэгили опубликовал работу, в которой изобразил некие тельца, возникающие на месте ядра во время деления клетки при образовании пыльцы у лилии (Lilium tigrinum) и традесканции (Tradescantia). Однако его рисунки не позволяют однозначно утверждать, что Карл Нэгили видел именно хромосомы.
В том же 1872 году ботаник Эдмунд Руссов привел свои изображения деления клеток при образовании спор у папоротника из рода ужовник (Ophioglossum) и пыльцы лилии (Lilium bulbiferum). На его иллюстрациях легко узнать отдельные хромосомы и стадии деления.
Некоторые исследователи полагают, что первыми увидел хромосомы немецкий ботаник Вильгельм Фридрих Хофмайстер задолго до К.Нэгили и Э.Руссова, еще в 1848-1849 гг. При этом ни К.Нэгили, ни Э.Руссов, ни тем более В. Хофмейстер не осознавали значения того, что видели.
В разных статьях и книгах приоритет открытия хромосом отдают разным людям, но чаще всего годом открытия хромосом называют 1882 г., а их первооткрывателем — немецкого анатома Вальтера Флеминга. Однако справедливее было бы сказать, что он не открыл хромосомы, а в своей фундаментальной книге «Клеточное вещество, ядро и деление клетки» ("Zellsubstanz, Kern und Zelltheilung") собрал и упорядочил сведения о них, дополнив результатами собственных исследований (рис. 1).
 Рисунок из книги В.Флемминга, изображающий разные стадии деления клеток эпителия саламандры (W.Flemming. Zellsubstanz, Kern und Zelltheilung. 1882г.)
Рисунок из книги В.Флемминга, изображающий разные стадии деления клеток эпителия саламандры (W.Flemming. Zellsubstanz, Kern und Zelltheilung. 1882г.)
В книге он описал непрямое деление ядра и привел много детальных рисунков. Ввел термины хроматин и митоз.
Термин «хромосома» был предложен немецким гистологом Х.Вальдейером в 1888г., «хромосома» в буквальном переводе означает «окрашенное тело», поскольку оснóвные красители хорошо связываются хромосомами.
Цитогенетика как наука сформировалась в начале 20 века. После переоткрытия в 1900г. законов Менделя потребовалось всего один-два года для того, чтобы стало ясно, что хромосомы ведут себя именно так, как это ожидалось от «частиц наследственности». В 1902г. Теодор Генрих Бовери (Германия) и в 1902-1903 гг. Уолтер Саттон и Эрнест Генри Вильсон (США) независимо друг от друга первыми выдвинули гипотезу о генетической роли хромосом. Они пришли к заключению, что именно хромосомы являются материальными носителями факторов наследственности, открытых Г. Менделем и позднее названных генами.
Т.Бовери обнаружил, что зародыш морского ежа Paracentrotus lividus может нормально развиваться только при наличии хотя бы одного, но полного набора хромосом. Также он установил, что разные хромосомы не идентичны по своему составу. У.Сеттон изучал гаметогенез у саранчового Brachystola magna и понял, что поведение хромосом в мейозе и при оплодотворении полностью объясняет закономерности расхождения менделевских факторов и образования их новых комбинаций.
В 1903 г. Сеттон в 1903 впервые использовал термин «цитогенетика».
В 1907 году американским исследователем К. Мак-Клонгом были открытыполовые хромосомы. Это событие было первым успехом применения микроскопа для решения задач генетики. Идентификация половых хромосом остается начальным этапом цитогенетического исследования всякого биологического объекта.
На начальных этапах развития изучались геномы, растений, низших животных, прокариот и вирусов. Цитогенетика человека и млекопитающих, занимающая ведущее место в современной цитогенетике, развилась позже, главным образом в связи с методическими трудностями.
Рассмотрим современные представления о макроструктуре хромосом. Прежде всего, необходимо отметить, что организация структур, отвечающих за наследственность у прокариотических и эукариотических организмов имеет отличия. В эукариотических клетках информация, необходимая для поддержания вида и нормального развития индивидуума, содержится в хромосомах, заключённых в клеточных ядрах, а у бактерий и сине-зеленых водорослей хромосомный материал, находится непосредственно в цитоплазме без ядерной оболочки.
Важная особенность хромосом заключается в их способности изменять свою структуру в зависимости от фазы клеточного цикла. Хромосомы как индивидуальные структуры становятся доступными для исследования после значительного укорочения и утолщения, которые они испытывают в период подготовки клетки к делению. Для соматических клеток таким делением является митоз, для генеративных — сначала митоз, а затем мейоз.
Хромосомы в этом состоянии представляют собой компактные палочковидные структуры разной длины с довольно постоянной толщиной, у большей части хромосом имеется перетяжка (первичная хромосомная перетяжка Х.п.), которая делит хромосому на два плеча. В области перетяжки расположена важная для удвоения хромосом структура, называемая центромерой (рис. 2).
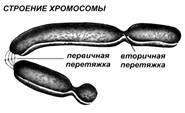
Рисунок 2.
Центромера – это особое образование, к которому прикрепляются нити веретена деления. Участки хромосомы, разделенные центромерой, называются плечами. Длинное плечо хромосомы обозначают буквой q, короткое — буквой p.
У некоторых хромосом имеются вторичные перетяжки - морфологический признак, позволяющий идентифицировать отдельные хромосомы в наборе. От первичной перетяжки отличаются отсутствием заметного угла между сегментами хромосомы. Вторичные перетяжки бывают короткими и длинными и локализуются в разных точках по длине хромосомы. Эти зоны называют зоны ядрышка (организаторы ядрышка). У человека вторичные перетяжки имеют 9, 13, 14, 15, 21 и 22 хромосомы.
Некоторые хромосомы имеют сателлит (спутник) — это округлое или удлинённое тельце, отделённое от основной части хромосомы тонкой хроматиновой нитью, по диаметру равный или несколько меньший хромосоме. Хромосомы, обладающие спутником принято обозначать SAT-хромосомами. Форма, величина спутника и связывающей его нити постоянны для каждой хромосомы.
Концевые зоны хромосом называются теломерами. У позвоночных теломеры состоят из богатых G повторов ДНК-последовательностей (TTAGGG) и специфических белков, создающих эти специализированные структуры. Взаимодействуя со многими другими факторами в клетке, теломеры способствуют динамичной регуляции поддержания стабильности хромосом.
В зависимости от морфологии выделяют 4-х типа хромосом:
- метацентрические (V-образные хромосомы, обладающие плечами равной длины);
- субметацентрические (с плечами неравной длины, напоминающие по форме букву L);
- акроцентрические- резко неравноплечие (палочковидные хромосомы с очень коротким, почти незаметным вторым плечом);
- телоцентрические (палочковидные хромосомы с центромерой, расположенной на проксимальном конце).
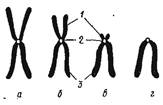
Рис. 3. Типы метафазных хромосом: а - метацентрическая; б - субметацентрическая; в - акроцентрическая; г- телоцентрическая: 1 - короткое плечо; 2 - центромера; 3 - длинное плечо
Хромосомы различаются не только по морфологии, но и по величине. Размеры хромосом растений и животных колеблются от долей микрона до десятков микрон (0,2-50). Средние длины метафазных хромосом человека лежат в пределах 1,5-10 микрон, диаметр от 0,2 мкм (или 200 А). Длина каждой определенной хромосомы относительно постоянна. Таким образом, каждая хромосома индивидуальна. Учитывая морфологию и величину хромосом, в клетке их можно точно идентифицировать, а для удобства изучения присваивать им определенные номера, что и было сделано для хромосом человека и некоторых других организмов.
Совокупность хромосом соматической клетки, определяемую их числом, величиной и формой, называют кариотипом. Хромосомы, несущие генетическую информацию, ответственную за развитие соматических признаков, называют аутосомами. В то время как последнюю пару хромосом, связанную с генетикой пола, называют половыми хромосомами или гоносомами. Для обозначения половых хромосом у различных видов используются различные символы (буквы), зависящие от специфики определения пола таксона (различные системы половых хромосом). Так, у большинства млекопитающих женский кариотип гомогаметен, а мужской гетерогаметен, соответственно, запись половых хромосом самки XX, самца — XY. У птиц же самки гетерогаметны, а самцы гомогаметны, то есть запись половых хромосом самки ZW, самца — ZZ. Например, нормальный кариотип женщины: 46, XX; нормальный кариотип мужчины 46, XY.
При проведении кариологических исследований кариотип систематизируют, т.е. располагают хромосомы по форме и размерам. Кроме диплоидного числа хромосом в кариотипе часто обозначают число хромосомных плеч. Для его определения подсчитывают число двуплечих хромосом (мета- и субметацентрических); иногда при определении Ч.х.п. двуплечими считают и телоцентрические хромосомы, применяя обозначение (NF). chromosome arm number, NF - число хромосомных плеч.
Для кариотипа используется запись в системе Международной цитогенетической номенклатуры (International System for Cytogenetic Nomenclature) ISCN 1995, имеющая следующий формат:
1.количество хромосом
2.половые хромосомы
3. особенности.
Положения хромосомной теории были развиты Т. Морганом и его школой. Согласно этой теории передача наследственной информации связана с хромосомами, в которых линейно, в определенной последовательности, локализованы гены. Основателями классической генетики являются Томас Хант Морган, Кэлвин Бриджес, Алфред Генри Стёртевант и Герман Джозеф Мёллер, чьи экспериментальные работы с плодовой мушкой (D. melanogaster) позволили сформулировать
основные положения хромосомной теории наследственности:
1.Кариотип вида постоянен (количественная, морфологическая, характеристика хромосом). Постоянство кариотипа обеспечивается механизмами митоза и мейоза;
2. Группы сцепления генов соответствуют определенным парам хромосом;
3. Индивидуальная структура хромосом постоянна, ее мутационные изменения наследуются;
4. Гены в хромосомах располагаются линейно. В ходе мейоза происходит рекомбинация между гомологичными хромосомами.
Эти выводы были опубликованы в 1915г. в книге «Механизмы Менделевского наследования» («The mechanisms of mendelian heredity»). В 1933г. за открытие роли хромосом в наследственности Т. Морган получил Нобелевскую премию по физиологии и медицине.
Положения хромосомной теории Т.Х. Моргана могут быть дополнены основными законами поведения хромосом у эукариот:
1. В кариотипе все хромосомы, исключая половые, парные, т.е. гомологичные. Половые хромосомы, в силу того, что они отличаются морфологически, называют гемилогичными;
2. Хромосомы каждой пары индивидуальны отличаются от других пар своими размерами и строением;
3. Хромосомы в ходе редукционного деления распределяются независимо, то есть при созревании половых клеток хромосомы разных пар распределяются в дочерние клетки независимо от других пар;
4. Закон постоянства числа хромосом: при митотических делениях сохраняется в дочерних ядрах то же число хромосом, что и в исходном ядре. При мейозе и образовании гамет число хромосом уменьшается вдвое, но после оплодотворения восстанавливается количество хромосом, характерное для данного вида.
В случае мейоза, диплоидный набор потомства состоит из пар гомологичных хромосом, имеющих одинаковые генный состав, размеры и структуру. Однако, этот набор хромосом уже смешанный, половина хромосом пришла от матери и вторая половина от отца. Дальнейшие процессы приводят к развитию зиготы в сложный многоклеточный организм. Весь процесс развития, при котором диплоидные клетки делятся и дифференцируются (образуют различные ткани и органы) представляет собой реализацию генетической и пространственной информации.
Если бы гаплоидный набор был совершенно одинаковым, то организмы несли бы идентичный набор хромосом и не отличались по этому набору от своих родителей. Тогда различия между рыбами сводилась бы исключительно к влиянию внешней среды (того водоёма), в котором они обитают. В действительности же картина более сложна.
В настоящее время выделяют три уровня организации наследственных структур у различных организмов (генный, хромосомный, геномный). Для понимания закономерностей наследственности исследования направлены по трем направлениям:
1) изучение морфологии и химического строения хромосом и кариотипа в целом;
2) выделение дискретных признаков организма, контролируемых единичными генами («инвентаризация» единиц наследственной изменчивости);
3) определение локализации генов в хромосомах (сцепление генов и карты хромосом). По каждому из этих разделов накоплено много данных, их интенсивная разработка продолжается как в теоретическом, так и прикладном (клиническом) аспектах.
Для понимания основных свойств и поведения хромосом необходимо изучение их тонкой структуры и химической природы.
Химический состав и микроструктура хромосом.
С химической точки зрения хромосома представляет собой нитевидную двуспиральную молекулу дезоксирибонуклеиновой кислоты (ДНК). Химический анализ хромосом показывает, что помимо ДНК, в хромосому включаются РНК и несколько низкомолекулярных основных белков – гистонов, а также и сложный кислый белок, называемый остаточным белком.
В хромосомах присутствуют также кальций, магний, железо и некоторые микроэлементы, роль которых ещё окончательно не выяснена. Можно в хромосоме найти и различные ферменты, в том числе ДНК - полимеразу, который участвует в репликации ДНК. ДНК-полимераза активируется ионами магния и возможно ионами марганца. Комплекс ДНК - гистон является основной структурной единицей хромосомы.
Основной вопрос заключался в том, каким образом молекула ДНК, достигающая в растянутом виде значительной длины упаковывается в процессе деления в компактные структуры - хромосомы?
Первоначально изучение структуры хромосом проводилось на делящихся клетках в профазе, так как тонкие хромосомы начинают в этот период утолщаться, конденсироваться. В ранней профазе хромосомы имеют вид тонких двойных нитей, которые получили название сестринские хроматиды. В метафазе они представлены в виде укороченных и утолщенных образований и в световом микроскопе видно, что хромосомы состоят из 4 нитей, которые были названы полухроматидами (хромонемами). Поэтому первоначально существовала гипотеза, по которой каждая хромосома состоит из многих нитей - хромонем (многонитчатая модель хромосомы). Согласно этой модели, общее число нитей в хромосоме — 64. Диаметр тончайшей нити составляет около 30 А. Многонитчатая структура хромосомы представлена на рисунке 5.
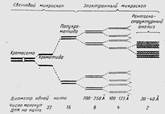
Рисунок 5. Схема микроскопической, субмикроскопической и молекулярной организации хромосомы (многонитчатая модель)
Другая гипотеза, которая подтвердилась в настоящее время, предполагала, что хромосома состоит только из одной нити, которая по мере подготовки к митозу претерпевает процесс спирализации. В ходе профазы спирализация распространяется по всей хромосоме, достигая максимума в метафазе. Поэтому в метафазе хромосомы выглядят очень компактными.
Значительная толщина хромосомы (диаметр 1400 нм) на стадии метафазы позволяет, наконец, увидеть её в световой микроскоп. Конденсированная хромосома имеет вид буквы X (часто с неравными плечами), поскольку две хроматиды, возникшие в результате репликации, по-прежнему соединены между собой в районе центромеры (рис. 6).
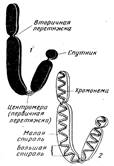
Рисунок 6. Схема строения метафазной хромосомы: 1 — морфология; 2 — внутренняя структура хроматиды, видимая при использовании специальных методов ослабления спирализации.
В телофазе наступает деспирализация хромонем, и в интерфазе хромонемы оказываются максимально раскрученными. Характер спирализации и деспирализации хромосом в митотическом цикле представляет закономерный процесс (цикл спирализации, рис. 7).
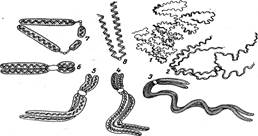
Рисунок 7. Схема спирализации хромонем в митотическом цикле: 1 — интерфаза, хромонемы слабо спирализованы (остаточные спирали); 2, 3, 4 — профаза, усиление спирализации хромонем, образование двух хроматид; 5 — прометафаза, проявление четырех полухроматид; 5 — метафаза, максимальная спирализация, выявляются как большая, так и малая спираль; 7 — анафаза; 8 — телофаза (одна из дочерних хромосом), деспирализация хромонем.
Со степенью спирализации хромосом связано их дифференциальное окрашивание по длине. При фиксации и окраске основными красителями разные участки (районы) дают разную реакцию. Одни участки интенсивно окрашиваются - их назвали гетерохроматиновыми. Гетерохроматиновые участки хромосомы называются хромомеры. Положение их в каждой хромосоме постоянно, а в разных хромосомах - различно.
Другие участки хромосом окрашиваются слабо — они названы эухроматиновыми. Эухроматиновые участки в интерфазе деспирализуются, что указывает на их более высокую метаболическую активность, т.е. глыбки хроматина в интерфазном ядре представляют собой не что иное, как гетерохроматиновые участки хромосом.
В хромосомах гетерохроматин располагается вне области центромеры. Половые хромосомы состоят почти полностью из гетерохроматина. Гетерохроматиновые и эухроматиновые нити представляют нити диаметром 25 нм.
Белки, участвующие в спирализации хромосом – гистоны, являются основными белками, так как содержат большой процент (20-30%) положительно заряженных аминокислот – аргинина и лизина. Гистоны взаимодействуют с ДНК, которая имеет отрицательный заряд благодаря отрицательно заряженным фосфатным группам.
У всех высших растений и животных в спирализации ДНК участвует 5 белков-гистонов, имеющих обозначение Н1, Н2А, Н2В, Н3 и Н4. Гистоны отсутствуют только в сперматозоидах некоторых организмов. Их функцию выполняют основные белки – протамины. Содержание гистонов в клетках соответствует их молярному соотношению: 1 Н1 :2 Н2А : 2 Н2В : 2 Н3 : 2 Н4.
Гистоны отличаются по молекулярной массе и аминокислотному составу. Гистон Н1 богат лизином (29%), гистоны Н2а и Н2в богаты как лизином, так и аргинином, а гистоны Н3 и Н4 богаты аргинином.
Гистоны формируют основные структурные единицы хромосом - нуклеосомы. Четыре гистона Н2А, Н2В, НЗ и Н4 образуют октамерный белковый комплекс (Н2А, Н2В, НЗ, Н4)2, который называют «нуклеосомный кор» (от англ. nucleosome core). Молекула ДНК «накручивается» на поверхность гистонового октамера, совершая 1,75 оборота (около 146 пар нуклеотидов).
Отдельные нуклеосомы связывает линкерная ДНК. В среднем линкерная ДНК составляет 60 пар (от 15 до 100) нуклеотидных остатков. Молекулы гистона H1 связываются с ДНК в межнуклеосомных участках (линкерных последовательностях) и защищают эти участки от действия нуклеаз (рис. 8).
В ядре каждой клетки присутствует около 60 млн. молекул каждого типа гистонов, а общая масса гистонов примерно равна содержанию ДНК, т.е. количество ДНК и гистонов в хроматине эквивалентно.
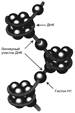
Рис. 8. Структура нуклеосом. Восемь молекул гистонов (Н2А, Н2В, НЗ, Н4)2 составляют ядро нуклеосомы, вокруг которого ДНК образует примерно 1,75 витка.
Аминокислотные остатки лизина, аргинина и концевые аминогруппы гистонов могут модифицироваться: ацетилироваться, фосфорилироваться, метилироваться или взаимодействовать с белком убиквитином (негистоновый белок). Модификации бывают обратимыми и необратимыми, они изменяют заряд и конформацию гистонов, а это влияет на взаимодействие гистонов между собой и с ДНК. Активность ферментов, ответственных за модификации, регулируется и зависит от стадии клеточного цикла. Модификации делают возможными конформационные перестройки хроматина.
В зависимости от степени спирализации и ее формы различают разные формы хромосом, в зависимости от степени конденсации. Одни хромосомы подвергаются циклу спирализации и деспирализации равномерно по всей длине на протяжении клеточного цикла. Другие - преимущественно состоят из гетерохроматина в течение всего жизненного цикла клетки, в том числе и в интерфазе, находятся в сильно спирализованном состоянии, а потому сильнее окрашиваются. Морфологию таких хромосом можно изучать в течение всего жизненного цикла клетки, а не только в период ее деления.
Например, дифференциация хромосом по длине хорошо видна на гигантских хромосомах. Гигантские хромосомы в 100-200 раз длиннее обычных и содержат в 1000 раз больше хромонем, чем обычные метафазные хромосомы большинства соматических и половых клеток.
Впервые гигантские (политенные) хромосомы были обнаружены Е. Бальбиани в 1881 г. в слюнных железах личинок мотыля (сем. Chironomidae). В дальнейшем оказалось, что такая структура хромосом характерна для ядер ряда соматических клеток личинок двукрылых - клеток кишечника, мальпигиевых сосудов, слюнных желез, а также найдена у некоторых растений (в антиподах и синергидах) и у простейших.
Гигантские хромосомы возникают при политении. Политения – процесс, при котором репродукция хромосом происходит без увеличения их числа в клетке. В этом случае 2 хромонемы после девяти последовательных удвоений образуют около 1000 нитей, плотно прилегающих друг к другу. Хромонемы гигантских хромосом постоянно находятся в частично и неравномерно деспирализованном состоянии, что обусловливает увеличение длины хромосом в 100-200 раз. Типичные гигантские хромосомы можно наблюдать в слюнных железах личинок дрозофилы (род Drosophila) (рис. 9, а). Строение и морфологические особенности этих хромосом видны в клетках при малом увеличении даже без специальной обработки на временных тотальных препаратах, но особенно хорошо - на окрашенных ацетокармином.
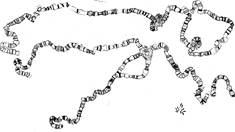
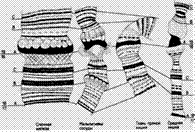
Рисунок 9. Относительные размеры хромосом в ядрах клеток слюнных желез (гигантские) и в клетках ганглия (митотические) дрозофилы; Примеры политенных хромосом
Например, если в любой соматической клетке хирономуса можно сосчитать 8 хромосом, то в клетке слюнной железы их только 4, так как одинаковые по морфологии и размеру хромосомы (одна отцовская, а другая материнская) обладают способностью объединяться, конъюгировать (соматическая конъюгация), что увеличивает еще больше толщину гигантских хромосом.
Другая особенность политенных хромосом состоит в том, что хромомеры многочисленных хромонем, плотно прилегая друг к другу, создают утолщения — диски, которые при окрашивании бывают более темными. Диск представляет собой участок плотно сложенной хроматиновой нити (хромомера). В световом микроскопе они выглядят в виде лент, поперечно исчерченных из-за чередования по всей длине интенсивно окрашенных участков (дисков) и светлых (междисковых) пространств (рис. 9, б). Хромомера политенной хромосомы содержит один или более генов в неактивном состоянии.
Размер и морфология дисков сильно варьируют, но для каждой хромосомы они постоянны и служат прекрасными маркерами при распознавании - идентификации хромосом (рис. 10). Между дисками хорошо видна политенность хромосомы.
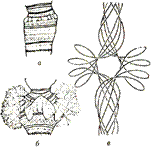
Рисунок № 10. Участок политенной хромосомы с дисковой стрктурой(а) и образованием пуфа (б).. Схема иллюстрирует возникновение пуфа путем деконденденсации четырех хроматиновых нитей, уложенных в хромомере (в).
Строение дисков изменяется в онтогенезе, что связано с функционированием хромосом. Так наблюдается попеременное набухание и разрыхление дисков — образование так называемых пуфов (рис. 10, б). Гигантские пуфы некоторых специфических дисков названы кольцами Бальбиани. Процесс образования пуфов представляет собой деконденсацию хроматиновых нитей, упакованных в диске (рис. 10, в) и является обратимым. В цитогенетике появление пуфов рассматривается в качестве морфологического выражения транскрипционной активности генов.
Другой моделью, на которой можно познакомиться с тонким строением хромосом и их функционированием, являются хромосомы типа «ламповых щеток». Вид этих хромосом действительно напоминает ершик, которым моют стеклянные пробирки. Отдельные участки этих хромосом сильно вытянуты и образуют симметричные петли, перпендикулярные оси хромосомы (рис. 11). Такое состояние хромосом встречается в ооцитах рыб, амфибий, рептилий и птиц.
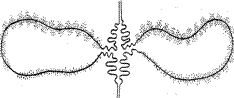
Рисунок 11. Схема строения отдельной петли, хромосомы типа «ламповых щеток».
В отличие от гигантских хромосом «ламповые щетки» не являются политенными, а содержат сильно деспирализованные хромонемы. Предполагают, что большая степень деспирализации связана с повышением метаболической активности хромосом в процессе роста ооцитов.
Тонкий электронномикроскопический анализ показал, что каждая хромонема по оси образует серию хромомер, из которых и выходят боковые петли - деспирализованные хромонемы, толщина их в самых тонких участках оказывается равной 100-200 А.
Характеристика кариотипов
Каждому виду организмов свойствен определенный кариотип. Кариотипы организмов могут содержать от 2-х хромосом у малярийного плазмодия до 1000 - у радиолярий. Примеры кариотипов приведены на рисунке 4.

Рисунок 4. Кариотипы разных видов растений и животных, изображенные в одном масштабе: 1 — диатомовая водоросль (Сосconcis placenttila); 2 — муха (Drosophila melanogaster); 3 — сложноцветное (Crepis capillaris); 4 — саранчовое (Gomphocerus rufus); 5 — жук (Gerris lateralis).
При изучении кариотипа парные (гомологичные) хромосомы располагают рядом. Примеры диплоидного числа хромосом у некоторых животных и растений приведены в таблице 1-2.
Таблица 1. Животные
| Plasmodium malariae | малярийный плазмодий | |
| Hydra vulgaris | гидра пресноводная | |
| Lumbricus terrestris | дождевой червь | |
| Bombyx niori | тутовый шелкопряд | |
| Pieris brassicae | капустная белянка | |
| Cyprinus carpio | Сазан | |
| Perca fluviatilis | Окунь | |
| Triturus vulga | Тритон | |
| Lacerta agili | ящерица прыткая | |
| Columba livia | Голубь | |
| Callus gallus | курица домашняя | |
| Lepus cuniculus | Кролик | |
| Bos taurus | крупный рогатый скот | |
| Anthropopithecus sp. | Шимпанзе | |
| Homo sapiens | Человек |
Таблица 2. Растения
| Abies, Picea, Pinus, Larix | пихта, ель, сосна, лиственница | |
| Cucumis sativus | Огурец | |
| Ribes rubruni | красная смородина | |
| Mains silvestris | Яблоня | 34, 51 |
| Quercus robur | дуб обыкновенный | |
| Soianum tuberosum | Картофель | |
| Secale cereale | рожь | 14, 28 |
Если сравнивать число и размер хромосом у человека и у других видов организмов, то можно увидеть огромные отличия. Например, у коровы, размер генома которой примерно равен геному человека, имеется 60 пар хромосом. У шпорцевой лягушки содержится всего 18 хромосом, но даже самые маленькие из них больше, чем самые крупные хромосомы человека. У птиц, наоборот, число хромосом достигает 40 и более и все они очень небольшие по размерам. Таким образом, количество хромосом в кариотипе не связано с уровнем организации животных и растений: примитивные формы могут иметь большее число хромосом, чем высокоорганизованные, и наоборот. Сравниваются кариотипы не только по числу хромосом, но и по массе ДНК. Применяемая единица измерения – пикограмм. 1 пг = 10 -12 г.
Считается, число и морфология хромосом в отдельных случаях могут служить показателем филогенетического родства видов. На этом принципе строится кариосистематика.
Хотя, сходство кариотипа указывает на родственные связи организмов, однако, и в пределах вида генетический материал претерпевает изменения (например, перекомбинации при мейозе), поэтому потомство отличается от родителей, хотя и несёт только генетические особенности, заложенные в хромосомах родителей.
Разнообразие кариотипов связано также с тем, что у некоторых организмов имеются добавочные к диплоидному набору хромосомы. Так, у ряда животных помимо крупных хромосом обнаружены очень мелкие «точечные» хромосомы. В отличие от хромосом нормального диплоидного набора, которые принято называть А-хромосомы, добавочные хромосомы назвали В-хромосомы. В-хромосомы часто называют необязательными, и долгое время данный тип хромосом оставался загадкой. В настоящее время В-хромосомы обнаружены у 80 видов животных и 256 видов растений (кукуруза, рожь, пресноводные тубеллярии, некоторые насекомые).
Впервые, В-хромосомы были обнаружены Эрнестом Вильсоном в 1905 г. у клопа Matepodius terminalis.
Свойства В-хромосом:
1. По размерам меньше хромосом основного набора;
2. Более интенсивно окрашиваются, т.к. имеют больше хроматина;
3. Присутствие не обязательно; наличие (отсутствие) в кариотипе не сказывается на фенотипе (однако их накопление в количестве больше 10, может вызывать депрессию роста, снижение плодовитости, различные аномалии в свойствах и признаках);
4. Не гомологичны А-хромосомам, в анафазе мейоза часто не наблюдается их равномерного распределения, поэтому в дочерние клетки может попасть неравномерное их число, что вызывает изменчивость кариотипа по числу В-хромосом (у кукурузы Zea mays их количество в клетке может варьировать от 1 до 34.);
5. В-хромосомы не содержат никакого генетического материала, т.е. не имеют экспрессирующихся генов с существенными для жизни организма функциями;
6. Для некоторых видов растений характерно явление соматической элиминации, т.е. В-хромосомы, например у растений, могут неизменно обнаруживаться в микроспороцитах, но не в клетках корня;
7. Обладают способностью к увеличению своего количества в последующих поколениях при половом размножении;
8. Как правило, В-хромосомы встречаются у малохромосомных видов.
Другая разновидность хромосом обнаруживается в кариотипе самок высших животных, они получили название тельца Барра. Тельца Барра – это половой, или Х-хроматин (вторая, неактивная половая хромосома). Величина телец Барра около 1 мкм. Они прокрашиваются всеми основными ядерными красителями более интенсивно, чем остальные хроматиновые структуры ядра. Тельца Барра наблюдаются в интерфазных ядрах соматических клеток самок плацентарных, включая человека.
Локализация Х-хроматина в ядре различна. В большинстве тканей он находится на внутренней поверхности ядерной оболочки и может иметь треугольную, плоско-выпуклую, трапециевидную, U-образную или гантелевидную форму. В веретеновидных и палочковидных ядрах Х-хроматин располагается на одном из полюсов ядра. Реже Х-хроматин располагается на ядрышке или в нуклеоплазме, при этой локализации он обладает сферической формой и трудно отличим от других хромоцентров, имеющих такой же размер, но неспецифических для пола.
Дата добавления: 2017-09-19; просмотров: 2738;