Типы мутации кариотипа эукариот
Хотя мы и говорим о законе постоянства числа и формы хромосом в наборе клетки для каждого вида организмов, однако следует отметить, что это постоянство относительно. Клетки разных тканей даже одного организма в зависимости от выполняемой функции могут содержать разное число хромосом. Так, например, в клетках печени животных бывает большее, чем два, число наборов хромосом (т. е. 4n, 8n).
Некоторые виды растений могут быть представлены формами, отличающимися числом хромосом, кратным гаплоидному. Так, рожь (Secale cereale) может иметь 2n (14 хромосом) или 4n (28 хромосом). При этом все видовые признаки у обеих форм ржи сохраняются. Такое явление - увеличение числа хромосом в организмах одного биологического вида сверх диплоидного числа (2n), кратно гаплоидному числу хромосом: 3n, 4n и т. д. называется полиплоидией. Полиплоидия была открыта Э. Страстбургером (Германия) в 1910 году. Полиплоидия регулярно встречается у растений и редко у животных (только у холоднокровных).
Различают автополиплоидию (удвоение собственного набора хромосом) и аллополиплоидию — соединение в одном биологическом виде диплоидных хромосомных наборов двух видов-предшественников. Существует также анеуплоидия — некратное изменение числа хромосом: потеря или добавление в хромосомном наборе отдельных хромосом. Все типы изменения числа хромосом объединяются понятием геномные перестройки.
Помимо них существуют и играют важную роль в наследственности хромосомные перестройки. Различают следующие типы структурных изменений хромосом: транслокация - обмен участками в пределах одной хромосомы или между разными хромосомами; инверсия - перевертывание участков хромосом на 180°; дупликация - удвоение участков хромосом, инсерция - вставка в каком-либо участке нуклеотидной последовательности, делеция - потеря отдельных участков и дефишенси – выпадение концевого участка хромосом. В некоторых случаях хромосомные перестройки могут приводить к изменению числа хромосом или хромосомных плеч. Такие типы перестроек хромосом, центрические слияния и разделения, названы по имени цитогенетика К. Робертсона (Великобритания), который в 1916 году впервые их описал. Их называют робертсоновскими транслокациями.
Хромосомные аберрации обозначаются дополнительными символами: делеция обозначается del, инверсия -inv, дупликация - dup, транслокация - t, инсерция – ins, r – кольцевая хромосома.
У многих животных хромосомный аппарат изучен детально, однако, у рыб изучение цитологического строения хромосом отстаёт от других видов животных. Основная причина отставания в изучении кариотипов рыб заключается в том, что исследователи ещё не освоили многообразия форм этого класса и до настоящего времени ещё описываются десятки новых видов рыб ежегодно. По данным разных авторов, описано свыше 20 000 видов рыб, что превышает общее количество видов пресмыкающихся, земноводных, птиц и млекопитающих, вместе взятых. При этом, каждый вид несёт свой генетический материал, то есть характерный только ему набор генов и, следовательно, обладают своим уникальным кариотипом.
Невысокая степень изученности кариотипов рыб связана также с особенностями их среды обитания, недоступностью глубоководной ихтиофауны для экспериментальных исследований в широком смысле слова.
Многочисленность рыб с одной стороны усложняет изучение генетической структуры этого класса, а с другой - обеспечивает эволюционную лабильность этой группы. В настоящее время считается, что рыбы являются бурно развивающейся группой и отдельные этапы генетических изменений, например, у лососевых, прослеживаются в течение нескольких десятилетий.
В целом, рыбы отличаются большим разнообразием кариотипов. Диплоидное число хромосом варьирует у разных видов от 12 до 250.
Хромосомы удобнее всего звучать на стадиях метафазы и анафазы митоза. Для этой цели у рыб берутся кусочки половых желез, мазки крови, в которых необходимо найти делящиеся лимфоциты. Наиболее полные исследования хромосом проведены на семействах карповых и лососевых рыб, а также на американских семействах ушастых окуней и зубастых карпов.
Кариотип белуги, где встречаются все типы хромосом, представлен на (Рис. 12).
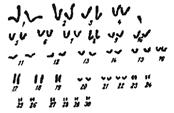 2n = 116±3 NF = (180 ±4) ±3
2n = 116±3 NF = (180 ±4) ±3
Рисунок 12. Хромосомный комплекс белуги, 1-30 пары хромосом.
Кариотип радужной форели представлен на рис. 13. Плечи хромосом соединены центромерой, участком, к которому прикрепляются нити веретена. В метафазе хромосома состоит из двух внешне одинаковых частей хроматид, а каждая хроматида состоит из 2-х тонких -нитей хромонем, которая деспирализуются после деления.

Рисунок 13. Метафазная пластинка (а) и кариотип (б) радужной форели (2n=60) (Уено, Миллер, 1971)
Число хромосом в кариотипах некоторых видов рыб представлены ниже:
Карповые:
- Серебряный карась 2n = 100
- Карп обыкновенный 2n = 100
- Карасекарп 2n = 100
- Возвратные гибриды 3n = 150
Осетровые:
- Осетр русский (многохромосомный вид – тетраплоиды) 2n = 247 ± 6
- Сибирский осетр (многохромосомный вид – тетраплоид) 2n = 238 ± 7, NF = (308 ±4) ±8
- Стерлядь (малохромосомный вид) 2n = 118 ± 3, NF = (176 ± 4)± 3
- Белуга (малохромосомный вид) 2n = 116±3 NF = (180 ±4) ±3
Гибриды:
- Бестер Бурцевский 2n = 117±3 NF = (174 ±7) ±3
(бестер товарный- 5-8 кг, 58 - метацентрических,
производство икры) 6 – акроцентрических, 54 ± 3 микрохромосомы
Кариотип бестера Бурцевского – базовая
цитогенетическая характеристика родительских видов: белуги и стерляди
- Бестер Аксайский 2n = 117±7 NF = 174 -175
(бестер стерляжий – 2-5 кг)
- Бестер Внировский2n = 117±1 NF = 176 -177
(бестер белужий – 15-30 кг)
Как уже отмечалось, диплоидное число хромосом у рыб сильно варьирует, но характер распределения по этому признаку показывает, что около 70 % видов имеют 42-50 хромосом (рис. 14).
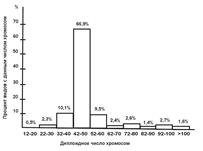
Рис. 14. Распределение рыб по числу хромосом (Кирпичникова, 1987)
Считается, что именно такие значения диплоидных чисел были характерны для предков современных рыб. В процессе эволюции наблюдались существенные преобразования кариотипов, которые приводили или к уменьшению, или к значительному увеличению числа хромосом. Наблюдаемую в настоящее время большую изменчивость по кариотипам объясняют тем, что рыбы представляют собой древнюю, очень гетерогенную группу животных, эволюция которой длится несколько сотен миллионов лет.
Рассмотрим более подробно механизмы мутаций, ведущих к изменению кариотипа у рыб.
У рыб, как и у других организмов, возможны различные типы мутаций. Генные, или точечные, мутации изменяют строение гена. Это может быть замена на определенном участке гена одного из нуклеотидов другим (например, гуанинового на тиминовый) в результате ошибки в момент репликации, выпадение одного или нескольких нуклеотидов из длинной цепочки ДНК или, наоборот, вставка нуклеотидов (и то и другое возможно при случайных разрывах цепи). Если при одном из таких изменений в середине гена образуется триплет (три нуклеотида) из числа бессмысленных кодонов, то на этом месте обрывается синтез белка и синтезируемая белковая молекула окажется укороченной, неспособной к нормальному функционированию. В остальных случаях меняется только порядок аминокислот в белке. Замена всего одной или двух аминокислот может оказаться по своим последствиям не очень существенной для жизнедеятельности особи, и мутация может сохраниться в популяции или даже может быть подхвачена отбором.
Данных о спонтанной частоте генных мутаций у рыб изучены не достаточно. Очевидно, она невелика: так, у карпа при просмотре 260 000 особей не удалось найти ни одной мутации генов чешуйного покрова (Цой и др., 1974б). Даже если принять, что в этом стаде возникла одна (незамеченная) мутация, частота мутирования составила бы всего 4 х 10-6. Мы можем судить о наличии мутационного процесса у рыб в природе только по косвенным наблюдениям. Об этом говорит наличие большой генетической изменчивости по морфологическим и физиологическим признакам в природных популяциях рыб.
Хромосомные перестройки составляют вторую большую группу мутаций. Как уже отмечалось, к их числу относятся транслокации, инверсии, дупликации и нехватки.
Транслокации, по-видимому, происходят достаточно часто, так даже близкие виды рыб отличаются по структуре кариотипа и эти отличия во многих случаях являются результатом эволюционного закрепления транслокаций. Вполне жизнеспособны индивиды с реципрокными транслокациями - взаимным обменом участками хромосом без потери или добавления генетического материала (рис. 15, а-в). Взаимные хромосомные транслокации имеют большое эволюционное значение и могут сопровождаться (хотя и не всегда) уменьшением или увеличением числа плеч. Иногда выживают и особи с добавочными участками хромосом.
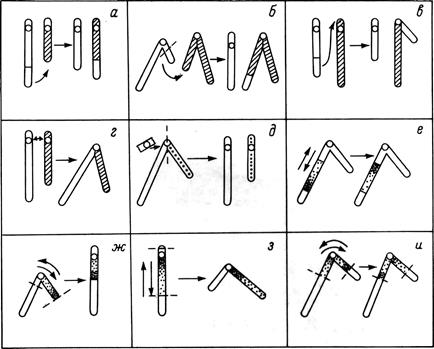
Рис. 15. Хромосомные перестройки (транслокации и инверсии) у рыб (схема): а - межхромосомная транслокация без изменения числа плеч; б - то же с уменьшением числа плеч; в - то же с увеличением числа плеч; г - центрическое слияние (fusion) робертсоновского типа с уменьшением числа хромосом; д - центрическое разделение (fission) с увеличением числа хромосом; е - неразличимая парацентрическая инверсия; ж - перицентрическая инверсия с уменьшением числа плеч; з - то же с увеличением числа плеч; и - неразличимая перицентрическая инверсия.
Еще более важны и, очевидно, довольно часты так называемые робертсоновские транслокации, или центрические слияния. Разрыв одной акроцентрической хромосомы происходит около центромеры, к месту разрыва присоединяется целая (или почти целая) другая хромосома, также акроцентрическая. Один или два незначительных околоцентромерных участка хромосом (с одной из центромер) теряются, и в результате две акроцентрические хромосомы превращаются в одну метацентрическую. Число плеч при этом остается неизменным (рис. 15, г). Реже происходит обратный процесс - центрическое разделение (рис. 15, д), для его осуществления нужна лишняя центромера. По недавним сообщениям, возможно и прямое деление центромеры на две дочерние (Imai, 1978).
Инверсии (рис. 15, е-и) могут быть отнесены к двум основным типам. Парацентрические инверсии, не захватывающие центральный участок, обнаружить трудно. Они возникают, вероятно, у рыб достаточно часто, но установить их наличие можно только при анализе наследования сцепленных генов. Перицентрические инверсии, включающие центромеру, широко распространены. Если два разрыва произошли в хромосомных плечах на равном расстоянии от центромеры, инверсию (без анализа маркерных генов) найти невозможно. При асимметричном расположении разрывов соотношение или даже число плеч в хромосомах изменяется.
Дупликации хромосомных участков у рыб безусловно происходят, хотя, вероятно, не часто. Наличие дуплицированных генов установлено чисто генетическими методами: под микроскопом дупликации заметить невозможно. Наиболее вероятным механизмом дупликации является неравный кроссинговер - обмен частями неточно конъюгировавших хромосом. Многие исследователи придают дупликациям особенно большое значение в эволюции рыб.
Нехватки, несомненно, также наблюдаются - и, вероятно, чаще, чем дупликации, - но подавляющее большинство нехваток резко снижает жизнеспособность их носителей и благодаря этому особи, несущие такие нехватки, быстро выбрасываются из популяций.
Спонтанная частота возникновения хромосомных перестроек у рыб неизвестна.
Третью группу мутаций составляют изменения плоидности - появление у нормально диплоидных видов рыб (2n) особей с уменьшенным вдвое (гаплоидным) кариотипом (n) или, наоборот, с увеличенными наборами - триплоидным (3n), тетраплоидным (4n) и другими, а также с увеличенным или уменьшенным числом отдельных хромосом (анэуплоидия).
Гаплоиды у рыб нежизнеспособны. При стимуляции развития яйцеклеток спермиями с предварительно разрушенным ядром почти все начавшие развиваться зародыши оказываются гаплоидными, развитие сопровождается уродствами и заканчивается гибелью эмбрионов в конце эмбриогенеза.
Чаще всего появляются триплоиды. По-видимому, у всех рыб с довольно большой частотой образуются диплоидные гаметы. Основной причиной их появления у самок служит слияние ядер яйцеклетки и второго направительного тельца; редукции числа материнских хромосом в этом случае не происходит. Слияние такой яйцеклетки с нормальным спермием (а также проникновение в нормальную яйцеклетку диплоидного спермия или двух спермиев одновременно - полиспермия) приводит к возникновению триплоидов. Они могут быть вполне жизнеспособными; так, триплоидами являются некоторые разновидности серебряного карася Carassius auratus gibelio (рис. 16) и отдельные расы живородящих рыбок Poeciliopsis и Poecilia.

Рис. 16. Триплоидия у серебряного карася Carassius auratus gibelio (no: Kobayashi et al., 1970). A - диплоидный набор хромосом (2n=100); Б - триплоидный набор (2n = 156).
Триплоидные особи найдены у радужной форели, у калифорнийской карповой рыбы Hesperoleucus symmetricus и у щиповок Cobitis taenia. Пониженная плодовитость триплоидов у двуполых рыб (связанная с неправильным распределением хромосом в мейозе), а иногда и полное их бесплодие являются, очевидно, причиной их отсутствия в популяциях большинства видов. Триплоиды нередко появляются при отдаленной гибридизации рыб. Тетраплоиды, вероятно, изредка также появляются в результате слияния диплоидных гамет, но о частоте их возникновения в природе никаких сведений нет.
Возможность анэуплоидии у рыб еще недавно ставилась под сомнение. Обнаружение особи с 85 хромосомами у гольца (2n =84), в том числе с тремя хромосомами, несущими маркерный ген Ldh-В2, позволяет предположить, трисомики могут в отдельных случаях выживать, во всяком случае у рыб, прошедших в прошлом через полиплоидизацию генома (лососевые, чукучановые и др.).
Ключевые слова и понятия:
А- и В-хромосомы, аутосомы, гоносомы, тельца Барра, кариотип;
Диплоидное и гаплоидное число хромосом, хроматиды, хромонемы, центромера, спутничные хромосомы, плечо хромосомы, теломера;
Метацентрические, субметацентрические, акроцентрические, телоцентрические хромосомы;
Гомологичные хромосомы, гемилогичные хромосомы;
Полиплоидия, автополиплоидия, аллоплоидия, анеуплоидия, вторичная диплоидизация;
Политения, политенные хромосомы;
Хромосомы «типа ламповых щеток»;
Дата добавления: 2017-09-19; просмотров: 907;