Диплоидное число хромосом
Рис. 1. Распределение видов по числу хромосом в различных таксонах рыб. а - Cyprinidae; б - Characidae; в - Lebiasinidae (Cypriniformes); г — Cyprinodontiformes (карпозубообразные). Полиплоидные виды исключены; для полиморфных видов указано максимальное из встречающихся число хромосом.
Интересно, что в некоторых группах карповых рыб, в частности у американских представителей рода Notropis, константность хромосомных наборов наблюдается, несмотря на значительную морфоэкологическую дивергенцию видов. Именно в этой процветающей группе рыб могли сохраниться полиплоидные мутанты, получавшие преимущество в конкурентной борьбе с обычными диплоидными формами. Полиплоиды возникли в трех семействах - Cyprinidae, Cobitidae (Вьюновые) и Catostomidae (Чукучановые). Это был несомненно важный этап в прогрессивной эволюции костистых рыб третичного периода. Среди карповых полиплоидными являются многие виды (не менее семи) в подсем. Barbinae и два или три - в подсем. Cyprininae. Карп Cyprinus carpio, золотая рыбка Carassius auratus и двуполая форма серебряного карася С. auratus gibelio имеют удвоенные наборы хромосом (2n = 98-104), однополые популяции С. auratus gibelio - утроенные наборы (3n около 150). Диплоидные и триплоидные формы сосуществуют среди нескольких подвидов серебряного карася в Японии, а подвид С. a. langsdorfi представлен даже тремя формами, имеющими в наборе 100, 156 и 206 хромосом. У обыкновенного карася из Дуная С. carassius кариотип состоит из 50 хромосом; эти данные, однако, требуют проверки. В подсем. Barbinae крупные виды (В. barbus, В. brachicephalus, В. mertdlonalis, В. tauricus, Tor putitora и другие) являются тетраплоидами (по происхождению); большое число мелких форм (рыбы из рода Puntius, ранее относившиеся к роду Barbus, и другие) - диплоидами. В двух подсемействах карповых рыб полиплоидизация произошла, очевидно, независимо. У вьюновых (сем. Cobitidae) найдено, наряду с диплоидами, несколько многохромосомных видов - Botia macracantus, В. modesta, Mlsgurnus fossilis. Наряду с этим у целого ряда видов обнаружены диплоидно-тетраплоидные и даже диплоидно-триплоидно-тетраплоидные комплексы:
Cobitis bivae . . . . . наряду с обычными формами с 2n = 48 имеются отдельные расы
с 2n = 96 (Sezaki, Kobayashi, 1978; Kimizuka, Kobayashi, 1983);
С. taenia………. 48(50), 72-75, 86, 94, 96-100хромосом (Kobayashi, 1976; Ueno et
al., 1980; Васильев, Васильева, 1982);
Misgurnus anguillicaudatus…….48(50) и 100 хромосом (Raicu, Taisescu, 1972; Ojima,
Takai, 1979).
Размеры эритроцитов у многохромосомных видов увеличены, также у них повышено и содержание ДНК в ядре. Полиплоидное происхождение карпов, карасей и некоторых вьюновых рыб подтверждается данными об увеличенном количестве у них дуплицированных локусов.
У всех 14 исследованных видов чукучановых рыб (Catostomidaе) в кариотипе насчитывается около 100 хромосом, содержание ДНК на геном повышено. Полиплоидизация генома у чукучанов, а также и у некоторых карповых рыб произошла, очевидно, позднее чем в середине третичного периода, т. е. более 50 млн. лет назад. Полиплоидные вьюновые рыбы, по-видимому, значительно моложе.
В эволюции ряда семейств карпообразных рыб прослеживается довольно четко и тенденция к уменьшению хромосомных наборов. Так, в семействе лебиасовых (Lebiasinidae) число хромосом у некоторых видов уменьшено до 22-30 (см. рис. 1). В этом случае связь между числом хромосом и специализацией более ясна, чем у лососевых.
В некоторых семействах (Characidae, Anostomidae и др.) многие виды имеют увеличенное число хромосом (более 50), при этом нередко хромосомы представляют собою двуплечие метацентрические элементы. При росте числа хромосом у харацид возрастало и количество ДНК. Таким образом, в отряде карпообразных эволюция кариотипа происходила с разной скоростью и в различных направлениях, и это привело к большой дивергенции хромосомных наборов.
Очень гетерогенным по кариотипам является отряд сомообразных (Siluriformes). У многих видов диплоидное число хромосом превышает 54-60, а некоторые многохромосомные виды сем. Сlаriidаe и Callichthyidae (Панцирные сомы) есть основания считать полиплоидами. Так, в роде Coridorus (Callichthyidae) имеется ряд видов с кратным увеличением числа хромосом и числа плеч.
| Число хромосом, 2n | Число хромосом, 2n | ДНК/n, (пг) | |
| Coridorus arcuatus, С. axelrodi и др. | 4.1 (1 вид)* | ||
| С. melanistus | - | ||
| С. myersi | - | 2,3 | |
| С. metae | - | ||
| С. julii | 4,2 | ||
| С. aeneus | 4,4 |
* Возможна ошибка в видовом диагнозе (Scheel, устн. сообщ.).
Содержание ДНК оказалось повышенным у видов с увеличенным числом хромосом.
Род Noturus (сем. Ictaluridae – Кошачьи сомы) по числу хромосом очень изменчив, диплоидные наборы у более чем 20 видов варьируют в пределах от 54 до 40 хромосом. Предполагается, что уменьшение числа хромосом происходило путем центрических слияний, перицентрических инверсий и других перестроек независимо и параллельно в двух филетических ветвях этого рода. Род Ictalurus в этом же семействе, наоборот, характеризуется большими хромосомными наборами (от 48 до 62).
Обширный и хорошо изученный отряд карпозубообразных (Cyprinodontiformes) может служить ярким примером связи между числом хромосом и специализацией. Предполагаемое исходное число хромосом (48) в сем. Cyprinodontidae сохранилось только у 45 % видов, у остальных оно уменьшено вплоть до наборов с 18-20 хромосомами (см. рис. 1). Сходная картина наблюдается и в сем. Гудеевые (Goodeidae). Среди живородящих Poeciliidae, наоборот, преобладает набор с 48 хромосомами.
В сем. Cyprinodontidae мы встречаемся с различиями в степени редукции числа хромосом между родами, с межвидовой и, наконец, внутривидовой изменчивостью кариотипов. В пределах многих политипических родов наблюдается одна и та же закономерность - наличие большего или меньшего числа видов с редуцированным набором. Приведем примеры такой изменчивости (по: Scheel, 1972).
| Род | Встречающиеся диплоидные числа у разных видов (в скобках - число видов) |
| Fundulus | 48(15); 46(4); 44(1); 40(2); 34(1); 32(1) |
| Rivulus | 48(5); 46(3); 44(2); 40(1) |
| Aplocheilus | 50 (4); 48 (7); 42 (1); 40 (2); 38(1), 34 (2) |
| Notobranchius | 44(1); 38(3); 36(3); 18(1) |
| Aphyosemion | 46(2); 42(4); 40(19); 38(4); 36(4); 34(3); 32(1); 30(3); 28(2); 22(2); 20(3); 18(1) |
По мере уменьшения числа элементов увеличивается количество крупных метацентриков (рис. 2); основным механизмом редукции надо считать центрические слияния. Большую роль играли и перицентрические инверсии.
У некоторых видов внутривидовая изменчивость кариотипов не уступает межвидовой. Так, для трех видов Aphyosemion получены следующие ряды кариотипов.
A. bivittatum 40, 38, 36, 34, 30, 26
A. calliurum 40, 38, 36, 34, 32, 30, 26, 22, 20
A. earneronense .... 34, 32, 30, 28, 26, 24
Вариация эта носит систематический характер, разновидности из разных мест имеют разное число хромосом. Анализируя вид A. bivittatum, Шеел отмечает, что кариотипы с наборами из 40 или 38 хромосом характерны для наиболее «генерализованных» разновидностей. В процессе заселения водоемов (миллионы лет назад) в обширных тропических лесах исходные формы должны были быть достаточно гибкими и, следовательно, иметь генотипы с максимальной изменчивостью (слабым сцеплением генов).

Рис. 2. Гаплоидные наборы хромосом различных рас Aphyosemion calliurum, схема (по: Scheel, 1972а). а - n =20; б - n = 17; в - n = 13; г - n = 10.
Впоследствии, после проникновения в те же маленькие, очень сильно изолированные водоемы других видов Aphyosemion, выгодной cтала максимальная специализация и ограничение изменчивости. В ходе отбора получали преимущество особи с малым числом хромосом, т. е. с более устойчивыми генными сочетаниями. Такой же процесс специализации и параллельного уменьшения числа хромосом происходил и при расселении A. calliurum. Сохранению каждого из видов способствовало наличие разновидностей с более пластичным генотипом, т. е. с меньшей степенью сцепления, с большим числом хромосом. При случайном вымирании сильно специализированных форм освободившиеся экологические ниши могут занимать эти достаточно пластичные разновидности.
Этот замечательный пример своеобразной адаптивной стратегии вида показывает, что у зубастых карпов в каждом отдельном случае эволюция кариотипа определялась равновесием нескольких сил. Главными из них являлись выгода специализации и в то же время необходимость сохранения достаточной пластичности. Излишне специализированные разновидности и виды, а может быть и роды, вымирали при неблагоприятных изменениях условий существования.
В одном из родов сем. Poeciliidae - Poeciliopsis - имеются триплоидные виды и разновидности с 72 хромосомами, возникшие, очевидно, в результате гибридизации и гиногенеза. Триплоиды могут появляться и при некоторых скрещиваниях видов из рода Poecilia.
Из остальных отрядов рыб лучше других исследованы некоторые семейства окунеобразных(Perciformes). В эту гетерогенную группу входят семейства с весьма различным уровнем пластичности и специализации. Широкая приспособляемость характерна, в частности, для окуневых (Percidae) и центрарховых (Centrarchidae), и мы видим, что в этих семействах почти все виды имеют 48 хромосом. Род Etheostoma (Этеостома, Дартеры) в семействе окуневых является исключением; многочисленные виды этого рода сильно специализированы, но у всех у них число хромосом остается неизменным (2n = 48). Вместе с тем такие семейства, как бычки (Gobiidae), хромисты (Cichlidae) и лабиринтовые (Anabantidae), включают много специализированных форм, и в этих семействах мы находим довольно много случаев численного уменьшения кариотипов.
Об отрядах скорпенообразных, камбалообразных, иглобрюхообразных и некоторых других таксонах наиболее продвинутых костистых рыб сказать что-нибудь трудно из-за малочисленности наблюдений. Отметим лишь, что у рыб всех этих отрядов, а также у колюшек, тенденция к уменьшению числа хромосом проявляется очень отчетливо.
Таким образом, сопоставление всех до сих пор исследованных рыбообразных и рыб позволяет сделать вывод о несомненном уменьшении как числа хромосом, так и количества ДНК на геном по мере продвижения от примитивных к более высокоорганизованным группам. Среди наиболее примитивных групп небольшие хромосомные наборы имеют миксины и двоякодышащие рыбы, но для них характерен высокий уровень содержания ДНК. В разных таксонах рыб наблюдается и увеличение и уменьшение числа хромосом. Редукцию хромосом, очевидно, нельзя приписать случайной фиксации робертсоновских транслокаций - более вероятным является отбор, диктуемый условиями существования той или другой группы и характером приспособлений рыб к окружающей среде. Уменьшение числа хромосом в ряде случаев связано, несомненно, со специализацией видов, требующей ограничения комбинаций генетического материала. Эта связь, однако, не абсолютна: изменение числа хромосом и количества ДНК в ядре может быть результатом приспособления к специфическим условиям существования и, в частности, необходимости изменения уровня метаболизма.
Главную роль в эволюции кариотипа рыб играли хромосомные перестройки типа центрических слияний (и разделений) и перицентрических инверсий. Можно предполагать, по аналогии с другими организмами, что в этом процессе большое участие принимают парацентрические инверсии, тандемные дупликации и мелкие нехватки, но пока мы не располагаем адекватными методами их обнаружения у рыб.
Полиплоидия неоднократно имела место в эволюции рыб. Доказанными можно считать случаи независимого возникновения полиплоидов в семействе осетровых, отрядах лососеобразных и карпообразных рыб и в меньшей степени в отряде сомообразных. Не исключено, что полиплоидия имела место и при эволюционном развитии некоторых других таксонов. Возможность выживания полиплоидов у рыб обусловлена наличием у многих из них сравнительно простого типа генетического определения пола, а у некоторых и полного отсутствия половых хромосом.
Эволюция хромосомного аппарата у двоякодышащих рыб представляет совершенно исключительный пример многократного увеличения хромосомного материала и параллельного увеличения размеров клеток. Этот пример может оказаться полезным при исследовании путей проникновения водных организмов на сушу.
Имеются указания на существование различий в числе хромосом между пресноводными и морскими рыбами, а также между глубоководными и прибрежными формами (Никольский, 1973; Никольский, Васильев, 1973). Предположение об увеличенном числе хромосом у пресноводных рыб вызывает большие сомнения, в целом ряде отрядов или в отдельных семействах пресноводные виды характеризуются сильно редуцированным числом хромосом, морские и проходные, наоборот, увеличенным. Ярким примером наличия редуцированных наборов могут служить, в частности, пресноводные зубастые карпы. Меньше сомнений вызывают данные о большем числе хромосом у арктических рыб, но и в этом случае не следует забывать о главном факторе дивергенции кариотипов рыб - степени их специализации, несомненно большей у обитателей тропиков и субтропиков.
В работе В. П. Васильев (1983) на большом статистическом материале показал наличие существенной положительной корреляции между кариологической изменчивостью внутри определенного таксона и степенью изоляции видов. Интересна и обнаруженная им отрицательная корреляция между кариотипическим разнообразием и плодовитостью рыб. Таким образом, дивергенция кариотипов рыб усиливается при наличии изоляции и сниженной плодовитости.
Дополнительно
Пример внутривидовой изменчивости по кариотипам можно рассмотреть на примере видового многообразия в подотряде Нототениевые.
В настоящее время известны хромосомные наборы 61 вида всех 8 семейств Notothenioidei, т.е. почти у половины (49%) всех видов подотряда. Изменчивость числа хромосом весьма существенна, о чем свидетельствует большой диапазон диплоидного числа хромосом от 2n=20 до 2n=58; NF=40–88.
В качестве предкового кариотипа у костистых рыб рассматривается кариотип с диплоидным числом хромосом равным 2n=48, при наличии только акроцентрических хромосом. В отряде Perciformes (Окунеобразных), к которому принадлежит подотряд Notothenioidei, число 2n=48 обнаружено у 73% всех кариологически исследованных видов. Предковый кариотип (2n=48, NF=48) среди нототениоидных рыб отмечен у 3 видов из двух разных семейств: Bovictidae - Щекороговые (C. gobio и B. angustifrons) и Bathydraconidae - Антаркти́ческие плосконо́сы, или батидра́ковые, (P. breviceps) (Природина 1986, 1990). Кариотипы этих видов одинаковы по числу хромосом и хромосомных плеч, что соответствует предковому состоянию кариотипа костистых рыб.
Такой кариотип, состоящий только из одноплечих элементов, можно назвать неспециализированным и недифференцированным. Более продвинутыми и специализированными считаются кариотипы, которые состоят из нескольких морфологических групп хромосом (мета-, субмета-, субтело- и акроцентрические хромосомы) и имеют число плеч более 48 (рис. 3).

Рис. 3. Кариограммы некоторых видов подотряда Notothenioidei: A – Cottoperca gobio Steindachner (Bovictidae)- Щекороговые, 2n=48, NF=48; B – Pleuragramma antarcticum Boulenger (Nototheniidae), 2n=48, NF=88: m – метацентрики, a – акроцентрики; C – Pseudotrematomus nicolai (Nototheniidae), 2n=58, NF=84; D – Prionodraco evansii Regan (Bathydraconidae), 2n=20, NF=40.
В целом, изменения кариотипа внутри подотряда Нототениевых шло в 3-х направлениях:
1. изменение морфологии хромосом без изменения их числа;
2. изменение количества хромосом в сторону уменьшения;
3. изменение количества хромосом в сторону увеличения.
Направление 1 представляет группа видов со стабильным числом хромосом 2n=48; кариотипы различаются только по морфологии хромосом, выражающейся в числе их плеч. К этому направлению специализации относятся 34 вида из 6 семейств: Eleginopidae - Патагониевые (1 вид), Pseudaphritidae - Конголлиевые (1 вид), Harpagiferidae - Антаркти́ческие рога́тки, или харпаги́феровые (2 вида), Channichthyidae - Белокровные рыбы, (13 видов), Nototheniidae (13 видов), Bathydraconidae- Батидраковые (4 вида). Внутри этой группы процесс кариологических преобразований (при постоянном числе хромосом) происходит в направлении увеличения числа двуплечих хромосом, а, следовательно, и числа хромосомных плеч от NF=50 до NF=88 (рис. 4).
Морфологические преобразования, вероятно, связаны с перицентрическими инверсиями, при которых участки хромосом занимают инвертированное положение, или процессом накопления генетического материала (гетерохроматина) на плечах хромосом.
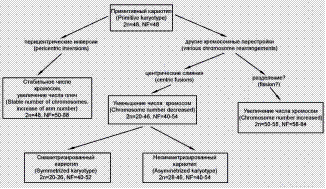
Рис. 4. Направления кариотипической специализации в подотряде Notothenioidei. (В.П. Природина)
Направление 2 связано с процессом увеличения числа хромосом, которое наблюдается у 3 видов из семейства Nototheniidae: 2n=50 (2 вида), 2n=58 (1 вид). Увеличение числа хромосом – довольно редкое явление, отмечено у рыб некоторых семейств карпообразных (Сypriniformes) (Кирпичников 1979). Это, вероятно, происходит за счет изменения хромосомной организации – центрических разделений хромосом.
Направление 3. Процесс уменьшения числа хромосом в сравнении с предковым кариотипом наблюдается у семейства Artedidraconidae - Борода́тковые (8 видов) и Nototheniidae (4 вида), 2n=46.
У видов, чья эволюционная судьба связана с дальнейшим уменьшением числа хромосом путем их центрического слияния, происходит элиминация хромосом. Кариотип, которому в процессе элиминации хромосом присуще наличие двуплечих и одноплечих хромосом, называется малохромосомным несимметризированным. Виды, имеющие такой кариотип, отмечены в семействах Nototheniidae (2 вида) и Bathydraconidae (2 вида) Они имеют хромосомные наборы 2n=28–38.
Дальнейшее уменьшение числа хромосом приводит к образованию малохромосомных кариотипов, состоящих только из двуплечих хромосом (2n=20–24) и имеющих число хромосомных плеч, равное удвоенному числу 2n, т.е. NF=40–48. Уменьшение числа хромосом происходит путем робертсоновских транслокаций, что приводит к образованию максимально симметризированного кариотипа, состоящего только из двуплечих элементов. К этой группе принадлежат 4 вида из семейства Nototheniidae и только 1 вид (Prionodraco evansi) из семейства Bathydraconidae, в кариотипе которого отмечено самое низкое число хромосом из изученных видов подотряда (2n=20, NF=40). Такое состояние всех хромосом следует рассматривать как «крайнюю» специализацию кариотипа, т.е. в этом случае процесс морфологической специализации кариотипа происходил путем центрических слияний, которые и обусловили уменьшение числа хромосом и образование метацентрических элементов в кариотипе. Кариотип, состоящий только из метацентриков, показывает завершенность эволюционных преобразований кариотипа, идущих путем центрических слияний. Уменьшенное число хромосом и их метацентрическое состояние у видов родов Notothenia, Pseudotrematomus -Трематомовые и Prionodraco - батидра́ковые показывает, что процесс эволюционных изменений кариотипа достиг своей максимальной продвинутости; что характеризует эти виды как высоко специализированные (рис. 5).
При сравнении всех направлений эволюционных преобразований кариотипа следует отметить, что в подотряде Notothenioidei наиболее распространены преобразования кариотипа, связанные с изменением морфологии хромосом, без изменения их числа. Такой тип эволюционных преобразований свойствен 77% общего числа кариологически изученных видов.
Таким образом, на примере подотр. Нототениевых видно, что большое таксономическое разнообразие сопряжено и с большим разнообразием кариотипов , как в семействах Nototheniidae и Bathydraconidae.
Рисунок 5. Антарктические плосконосы: Прионодрако Эванса (Prionodraco evansii). Антаркти́ческие плосконо́сы, или батидра́ковые, или плосконо́совые (лат. Bathydraconidae) — семейство морских автохтонных антарктических донных рыб
Дата добавления: 2017-09-19; просмотров: 1966;