Исходный материал для селекции лесных древесных пород
Под исходным материалом подразумевают культурные и дикие формы растений, используемые для выведения новых сортов. Основными источниками исходного материала являются естественные (местные и интродуцированные) популяции, гибридные популяции, полученные на основе внутривидовых и отдаленных скрещиваний, инцухт-линии, индуцированные мутанты и полиплоиды (Г.В. Гуляев, В.В. Мальченко, 1983).
Успех селекции обусловлен формовым разнообразием древесных растений. Лесные деревья настолько изменчивы, что почти невозможно найти два экземпляра, которые были бы полностью сходны по строению, внешнему виду и жизненным проявлениям (Э. Ромедер, Г. Шенбах, 1962). Чтобы ориентироваться в этом многообразии, ботаники делят растительный мир на классы, семейства, роды, виды, подвиды, разновидности и другие надвидовые и внутривидовые таксоны.
Причиной фенотипического разнообразия являются внешние (среда) и внутренние (наследственность) факторы. Разграничить влияние среды и наследственности на формирование конкретного фенотипа возможно только при постановке специальных опытов, в которых нормируется влияние среды или генотипа. Сила этих влияний могут быть самыми разными в зависимости от условий испытания, характера исследуемого материала и изменчивости изучаемых признаков. При этом различают изменчивость морфологических (форма ствола, кроны, корки; форма, длина, ширина, толщина и окраска хвои и листьев; цвет и форма цветков, шишек, семян, и т.п.), физиологических (фенологические признаки, рост и др.), качественных признаков, а также признаков устойчивости.
Многообразие форм является результатом непрерывного мутационного процесса, отдаленного внутри- и межвидового скрещивания и полиплоидизации. Степень полиморфизма зависит от возраста видов, а также условий их произрастания. Филогенетически молодые и эволюционно старые виды характеризуются, как правило, меньшей внутрипопуляционной изменчивостью по сравнению с видами, находящимися в расцвете своего развития. Гетерогенность популяций обычно значительна в области оптимума вида. В экстремальных условиях изменчивость деревьев по ряду признаков выражена слабее, наряду с этим частота отдельных форм здесь бывает повышенной.
За исключением случаев модификаций, связанных с влиянием биотических и абиотических факторов, полиморфизм популяций генетически обусловлен. Однако имеются значительные различия в степени генетического контроля тех или иных признаков. При этом выделяется группа константных, строго детерминированных признаков, не зависящих от внешних факторов, и группа лабильных признаков, изменяющихся в той или иной мере под влиянием среды. Характер изменчивости некоторых признаков у древесных растений зависит также от типа их онтогенеза.
Фенотипическое проявление нормы реакции особей в каждом случае находится в определенных пределах, что связано с функциями соответствующих органов, обеспечивающих индивидууму жизне- и конкурентоспособность в довольно широких диапазонах среды произрастания. В изменении отдельных групп признаков наблюдаются корреляции. Последние отмечаются, в частности, между некоторыми морфологическими признаками древесных растений и физико-техническими свойствами древесины, интенсивностью их роста, смолопродуктивностью, устойчивостью к биотическим и абиотическим факторам и другим свойствам, имеющим важное хозяйственное значение.
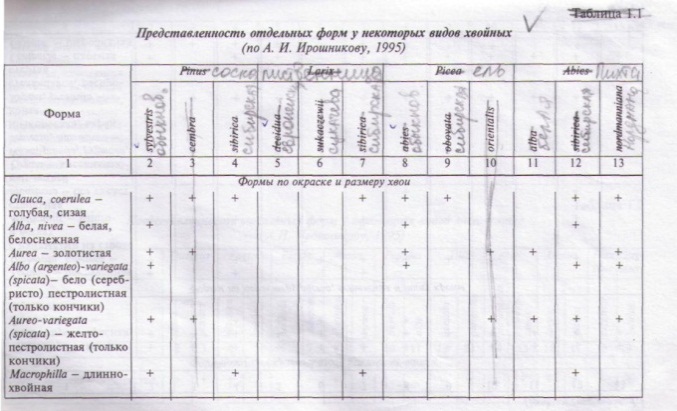
Многообразие форм древесных пород давно было подмечено ботаниками, садоводами и лесозаготовителями. На первом этапе дендрологов прежде всего интересовали резко уклоняющиеся формы с декоративными свойствами, представляющие особую ценность для зеленого строительства. В течение XVII-XIX веков в парках и ботанических садах многих стран мира коллекционировали деревья с пирамидальной, плакучей, колонновидной, зонтикообразной, шаровидной, веерообразной, шатровидной как плотной, так и ажурной формами кроны; с причудливыми ветвлениями и изгибами стволов; разнообразными наростами и наплывами на ветвях и стволе; гладкой, отслаивающейся или мощной коркой; с признаками гигантизма или, наоборот, карликовости; с кустовидной или стелющейся формой; с различными окраской, деформацией (рассеченность, скрученность) и опушением листьев; с яркой окраской или аномалиями в развитии, размере и форме цветков, микро- и мегастробилов, плодов, шишек и др. Изучение формового разнообразия стимулировали и работы Н.И. Вавилова по параллельной изменчивости у древесных растений (табл.1.1 и 1.2).
Из всего многообразия селекционеры отбирают ценные в каком-либо отношении фенотипы и разрабатывают меры по их сохранению. При этом предлагается: а) выделение лесных генетических резерватов; б) сохранение отдельных площадей (в особенности изолятов) насаждений и деревьев (эталонных, плюсовых); в) создание испытательных (коллекционных) культур и клоновых архивов; г) создание банков семян, пыльцы и меристем (С.А. Мамаев и др., 1988). Более подробно проблема сохранения генетического фонда лесных древесных пород в местах естественного (in situ) и вне мест естественного (ex situ) произрастания (плантации, коллекции, архивы, банки семян, меристем, пыльцы и т.п.) рассмотрена ранее (А.П.Царев и др., 2000).
***
К сожалению, в силу ряда причин не все из полученных и отмеченных в настоящей главе достижений нашли использование в производстве. Кроме того, в последние годы многие из направлений исследований пришлось свернуть из-за недостатка средств. Наблюдаются неблагоприятные тенденции и в других странах мира. Они заключаются в истощении невозобновляемых ресурсов, уменьшении биологического разнообразия, снижении доли лесопокрытой площади, вырубке наиболее производительных насаждений, развитии других неблагоприятных факторов, с одной стороны, и увеличении потребности в различных видах полезностей леса, с другой. Очевидно, что требуются какие-то меры по преодолению этих неблагоприятных тенденций. Необходимо наметить задачи ближайшего будущего (программа-минимум) и более отдаленного (программа-максимум).
Продолжение табл. 1.1
| Формы по окраске молодых женских шишек | ||||||||||||
| Chlorocarpa — зеленая Erithrocarpa — красная Bicolor, dichroa - | + + + + | + + + | + + + + | + + + | + + + | + + + | + + + | + + + | + + | + + + | + + + | + + |
| двухцветная | ||||||||||||
| Формы по характеру роста, ветвления и типу кроны | ||||||||||||
| Tortuosa — покрученная Gibberosa, tuberosa — | + + | + | + + | + | + + | + | + | + | + | |||
| с наплывами на стволе | ||||||||||||
| и ветвях Multicaulis — много- | + | + | + | + | + | + | + | + | + | + | ||
| ствольная Monstrosa — без скелет- | + | + | ||||||||||
| ных ветвей Virgata - змеевидная Nana, pumila, pygmaea, depressa, humistrata — | + | + | + | + | + | + | + + | + | + | + + | + | |
| низкорослая, карликовая Repens, prostrata -стелющаяся, распростертая | + | + | + | + | + | + | + | + | + | + | ||
| Compacta - плотная Globosa — шаровидная | + + | + | + + | + + | + + | + + | + | + | + | + | + |
| Окончание табл. 1.1 | ||||||||||||
| Pendulajnversa, vimi- | + | + | + | + | + | + | + | + | + | + | ||
| nalis, nutans — | ||||||||||||
| плакучая, свисающая, | ||||||||||||
| плетьевая, пониклая | ||||||||||||
| Fastigiata, pyramidalis, | + | + | + | + | + | + | + | + | + | + | + | + |
| cupressiana — узко- | ||||||||||||
| пирамидальная, кипа- | ||||||||||||
| рисовидная, веретено- | ||||||||||||
| образная | ||||||||||||
| Columnaris — колонно- | + | + | + | + | + | + | + | + | + | + | + | + |
| видная |
Таблица 1.2
Дата добавления: 2017-01-13; просмотров: 973;