Изменчивость живых организмов
Изменчивость — это процесс возникновения различий между особями по ряду признаков (размеры, форма, окраска, химический состав и пр.). Наряду с наследственностью и отбором она является основой органической эволюции.\ Более подробно явление изменчивости и методы ее изучения были уже рассмотрены авторами (2000). Здесь необходимо вспомнить основные положения, которые характерны для изменчивости, поскольку этот параметр является одним из важнейших для эффективной селекции.
Выделяют изменчивость качественную (олигогенную, альтернативную), основанную на изменчивости по качественным признакам, и количественную (полигенную, непрерывную, флуктуирующую), основанную на изменчивости по количественным признакам.
Качественные признаки контролируются, как правило, одним или немногими генами, действие которых четко отделено от действия ненаследственных факторов. Различия по качественным признакам устанавливаются непосредственно путем наблюдения или сравнения без применения измерения или взвешивания. Качественные признаки обладают прерывистой изменчивостью. Описываются они по принципу «есть — нет»: опушенный — неопушенный, окрашенный — бесцветный, трещиноватый — гладкий, пирамидальный — раскидистый. Большая часть качественных признаков при гибридизации наследуется в соответствии с законами Г. Менделя. Отсюда и наследственная качественная изменчивость обусловлена индивидуальным или комбинированным действием одной или нескольких пар аллелей, расщепление по которым подчиняется мен-делевским закономерностям.
Количественные признаки (мерные, полигенные) контролируются суммарным действием большого числа генов. Эти признаки могут быть определены путем измерения, взвешивания, подсчета. Вследствие полигенного контроля и большой модификационной изменчивости под влиянием условий внешней среды количественные признаки обладают непрерывной изменчивостью. Они могут быть описаны по принципу «больше — меньше», т.е. один и тот же признак, присущий разным особям или формам, имеет различную степень выражения. К количественным признакам относятся: диаметр и высота ствола, прирост и запас, угол прикрепления сучьев, плотность древесины, урожайность семян и т.п. Проявление количественных признаков в значительной степени зависит от условий внешней среды.
Изменчивость органически присуща всем живым существам. Немецкие лесоводы еще в XIX веке констатировали, что у каждого дерева свое лицо. Ботаники попытались упорядочить результаты этой изменчивости. Так, Н.И. Вавилов, В.Н. Сукачев и др. вслед за исследованиями Ч. Дарвина, Э. Цедербауэра и голландского ботаника Я.П. Лотси показали, что виды слагаются из большого числа биотипов (форм), которые отличаются как морфологическими, так и физиологическими особенностями. Было установлено, что виды могут быть крупными или мелкими. Первые Лотси называл «линне-онами» в честь К. Линнея, а вторые — «жорданонами» в честь французского ботаника Жордана, который первым попытался вычленить из установленного К. Линнеем вида крупки весенней (Draba verna L.) до 200 мелких наследственных форм, принимаемых им за виды. Среди систематиков до сих пор идут споры об объеме и отличительных признаках такой основной систематической категории, как вид. Однако постепенно на смену представлениям о неделимом монотипном виде приходит понятие о политипической структуре вида, сторонниками которой были и наши ученые В.Н. Сукачев и Н.И. Вавилов.
В.Н. Сукачев отмечал среди внутривидовых следующие категории:
• экотипы — наборы биотипов, отвечающие определенным условиям существования, включающие климатипы, эдафотипы, це-нотипы — как результат групповой изменчивости;
• лузусы — формы, отличающиеся какими-либо признаками, но 11С обладающие определенным ареалом или не относящиеся к опреде-испиым экотипам, например красно- и зелено-шишечные формы ели;
• аберрации — редкие резко уклоняющиеся формы: змеевидные, пнакучие и т. п.;
• экады — ненаследственные формы, или модификации.
Для практической селекции очень большое значение имеет закон гомологических рядов в наследственной изменчивости, сформулированный Н.И. Вавиловым в 20-е годы и доложенный в Саратове на III Всероссийском съезде селекционеров в июне 1920 г. Сущность закона заключается в том, что признаки близкородственных видов и даже родов изменяются параллельно. Например, плакучие и пирамидальные формы есть и у тополя черного, и у тополя белого. Более того, и у рода тополя в целом, и у рода ивы, и у других родов древесных растений. Таким образом, зная формовое разно образие одного вида или рода, можно прогнозировать его у родственных видов.
В последние десятилетия изменчивость лесных пород стали изучать на биохимическом уровне. При этом исследуются изозимы (ал-лозимы), антигены, терпены и другие вещества.
Изозимы — это множественные формы энзимов (специализированных белков, которые ускоряют биохимические реакции), синтез которых контролируется более чем одним геном, в то время как аллозимы — альтернативные формы фермента, кодируемые различными аллелями одного и того же гена. К настоящему времени для самой изученной в этом плане сосны обыкновенной исследовано 26 систем энзимов, для дуба — немногим более 10 (W. Prus-Glowacki, 1991; A. Zanetto et all., 1994; R. Bacilieri et all., 1995). К числу наиболее часто используемых систем энзимов относятся эстераза, глута-мат-оксалат-трансминаза, лейцин амино-пептидаза, кислая фосфа-таза, рибонуклеаза, алкоголь дегидрогеназа, малат дегидрогеназа, глутамат дегидрогеназа, шикимат дегидрогеназа, пептидаза, фос-фоглюкоизомераза и др. Исходят из того, что эти энзимы кодируются определенными локусами или аллелями. Общее число энзиматических локусов, описанных, например, для Pinus sylvestris, колеблется от 37 до 54, а число аллелей, кодированных в локусах, — от 90 до 110. Определение энзимов производят в крахмальном геле с помощью электрофореза. Аллели обозначаются номерами от катода к аноду.
По результатам электрофоретического анализа гомогената энзимов производят вычисления на видовом, внутри- и межпопуляционном и организменном уровнях. Учитывают число аллелей, генное разнообразие, эффективное число аллелей и др. Подсчитывается статистическое число аллелей на локус {Ар), наблюдаемая гетерозиготность (Нор), ожидаемая гетерозиготность (Hep) и эффективное число аллелей (Аер).
Как и все биохимические исследования, эти методы требуют узкой специализации персонала, недешевых лабораторного оснащения и расходных материалов. Поэтому они могут быть использованы только в немногих научных организациях. Более детальное их описание дано в (кроме уже цитированных) работах В.А. Духарева (1978); Г.Г. Гончаренко, В.Е. Падутова (1988); Н. Hattemer et al. (1993); A. Kremer, R.J. Petit (1993) и др.
Однако эти методы не всегда отражают реально существующее разнообразие и изменчивость признаков. Так, А.И. Ирошников (1996) отмечает контрастность характеристик межпопуляционной изменчивости видов с обширными ареалами (сосен обыкновенной и сибирской, лиственницы сибирской), полученных по материалам изоферментного анализа, с одной стороны, и результатами испытания потомств популяций в географических культурах, созданных в разных регионах, с другой стороны. Если в первом случае фиксируется как правило 3-5%-ный (0,5-12%-ный) уровень межпопуляционной генетической изменчивости, то во втором — он достигает 20-30% и более. У ряда популяций (как маргинальных, так и из оптимума произрастания вида) происходит полная элиминация потомства при его выращивании в контрастных условиях в пределах ареала. Очевидно, что требуются дальнейшие исследования как по уточнению методов, так и по их приближению к реально существующей изменчивости фенотипов в пределах ареалов видов.
Пионерные работы с антигенами у сосны обыкновенной были осуществлены в серологических опытах М. Хагманом(М. Hagman, 1967, 1977, цит. по W. Prus-Glowacki, 1991). Однако из-за трудоемкости этих методов они находят применение в основном при специфических исследованиях. Например, когда надо различить генотипы, которые другими методами различить не удается.
Терпены широко распространены в растениях. Так, у сосны обыкновенной идентифицировано 11 монотерпенных соединений: аль-фа-пинен, бета-пинен, мирцен, лимонен, бета-фелландрен, саби-нен, дельта-3-карен, трициклен, камфен, терпинолен и транс-бета-оцимен. Они использовались в качестве генетических маркеров при исследовании генетического состава популяций лесных древесных пород (А.Е. Squillace, 1976, цит. по W. Prus-Glowacki, 1991; А.В. Чудный, 1979; А. И. Чернодубов, 1999 и др.).
В общем виде изменчивость можно выразить следующей формулой:
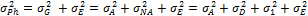
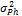 — наблюдаемая фенотипическая изменчивость;
— наблюдаемая фенотипическая изменчивость; 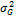 — генотипическая изменчивость;
— генотипическая изменчивость; 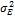 : — изменчивость, обусловленная средой;
: — изменчивость, обусловленная средой; 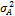 —аддитивная доля изменчивости в общей фенотипической изменчивости;
—аддитивная доля изменчивости в общей фенотипической изменчивости; 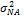 - неаддитивная доля изменчивости;
- неаддитивная доля изменчивости; 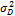 — доля изменчивости, обусловленная доминированием;
— доля изменчивости, обусловленная доминированием; 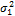 — доля изменчивости, обусловленная эпистазом.
— доля изменчивости, обусловленная эпистазом.
Кроме того, фенотипическая изменчивость некоторыми исследователями (С. А. Мамаев) делится на: внутривидовую и внутриор-ганизменную, или эндогенную.
Внутривидовая изменчивость подразделяется на:
• индивидуальную — проявление дифференциации особей в пределах вида;
• половую — зависящую от пола;
• хронографическую (временную), включающую возрастную (онтогенетическую) и сезонную изменчивости;
• экологическую, обусловленную воздействием на растение определенных факторов окружающей среды;
• географическую — результат дифференциации вида в пределах ареала в широтном и меридиональном направлениях;
• гибридогенную — на границах ареалов в местах спонтанной гибридизации.
Эндогенная (или метамерная) изменчивость — это изменчивость органов (листьев, цветков, плодов, семян и т. п.) в пределах особи. В зависимости от уровня изменчивости тех или иных видов оценивается их исходный материал. Изучение изменчивости исходных популяций позволяет выбрать наиболее эффективный метод селекции.
Дата добавления: 2017-01-13; просмотров: 1189;