Гормон роста гипофиза и инсулиноподобный фактор роста-1
ГР, также известный как соматотропин, представляет собой довольно крупный одноцепочечныый полипептид. Молекулярная масса гормона приблизительно 22 кДа, он состоит из 191 аминокислоты и имеет два дисульфидных мостика внутри цепи. Он образуется в соматотропных клетках, наиболее многочисленных клетках передней доли гипофиза (Rijnberk, 1995; Javorsky et al, 2011). Аминокислотная последовательность ГР собак идентична последовательности ГР свиней; кошачий ГР отличается от собачьего ГР только одной аминокислотой. ГР человека (и других приматов) отличается значительно (примерно на 33%) (Ascacio-Martinez and Barrera-Saldana, 1994; Castro-Peralta and Barrera-Saldafia, 1995; Warren et al 1996; Liu et al, 2001; Wallis, 2008).
У большинства видов млекопитающих ГР кодируется одним геном; у человека и других приматов группа из пяти генов кодирует ГР гипофиза и хорионный соматотропин (группа GH-hCs). Ген hGH-N кодирует белок 22 кДа (191 аминокислота), при этом транскрипция происходит избирательно в соматотропных клетках гипофиза. Другие гены экспрессируются в различных структурах плаценты. Известно, что у людей соматотропные клетки гипофиза секретируют смесь нескольких форм ГР, образующихся из формы массой 22 кДа в ходе посттрансляционной модификации. «Нормальный» белок массой 22 кДа из 191 аминокислоты представляет собой основную физиологическую форму и составляет 75% секретируемого гипофизом ГР (Melmed et al, 2011; Barret et al, 2012).
У млекопитающих ГР секретируется импульсами, как и другие гормоны передней доли гипофиза. Импульсы индуцируются главным образом эффектом рилизинг-гормона ГР (РГГР), в то время как минимумы секреции обусловлены в основном соматостатином (Melmed et al 2011). Число импульсов секреции ГР у здоровых собак незначительно различается по данным разных исследований: 1-3 импульса/6 ч, 1 импульс/4,5 ч, 2-7 импульсов/12 ч (Cowan et al, 1984; French et al, 1987; Beijerink et al, 2011). У людей наступление сна является сильным стимулом секреции ГР, однако у собак не обнаружено различий между фазами сна и бодрствования (French et al, 1987). У разных видов млекопитающих показаны возрастные и половые различия в секреции ГР. Тем не менее, секреция ГР одинакова у сук (во время анэструса) и кобелей; при этом старение у собак сопровождается снижением секреции ГР (Gobello et al, 2002; Lee, 2004). Секреция ГР у некастрированных сук изменяется во время лютеиновой фазы полового цикла, так как в периоды высокой концентрации прогестерона в плазме базальная секреция ГР выше, а импульсная секреция ГР – ниже. Это объясняется частичным подавлением секреции ГР гипофизом при синтезе ГР в молочных железах, индуцированном прогестероном (см. ниже) (Kooistra et al 2000b). Систематическая оценка характера секреции ГР у кошек пока не проводилась. В циркулирующей крови ГР в некоторой степени связывается с ГР-связывающими белками, что уменьшает колебания концентрации ГР и продлевает время полувыведения из плазмы. Рецептор ГР собак изучен на молекулярном уровне и его кодирующая последовательность в значительной степени гомологична последовательности рецептора ГР нескольких других видов. После связывания ГР рецептор димеризуется и активирует тирозинкиназы из семейства янус-киназ (ЯК) в цитозоле (van Garderen etal, 1999).
Изначально полагали, что эффект ГР обусловлен непосредственным действием гормона на ткани. В 1957 году Salmon and Daughaday открыли факторы роста, названные факторами сульфирования и в последующем переименованные в соматомедины Daughaday et al (1972). После того, как Rinderknecht and Humble (1978a, b) удалось выделить эти вещества, они получили название инсулиноподобных факторов роста (ИФР) – термин, использующийся до настоящего времени. Существует два основных ИФР: ИФР-1 (ранее соматомедин-С), который наиболее важен, и ИФР-2. Изначально предполагалось, что ГР индуцирует синтез ИФР-1 в печени, который затем действует на ткани-мишени и способствует их росту. В настоящее время предполагается, что ГР и ИФР-1 действуют как согласованно, так и независимо, стимулируя пути, ведущие к росту и регулирующие многие метаболические процессы. Кроме того, стало очевидно, что ИФР-1 образуется не только в печени, но и почти во всех тканях организма, где действует аутокринным/паракринным способом. Концентрация циркулирующего ИФР-1, происходящего в основном из печени, соответствует концентрации ГР (т. е. при состояниях, когда концентрация ГР низкая, концентрация ИФР-1 также снижается, и наоборот) (Le Roith et al, 2001). Заболевания печени также влияют на ИФР-1, так как его концентрация снижается с уменьшением функциональной массы печени (Styne, 2011).
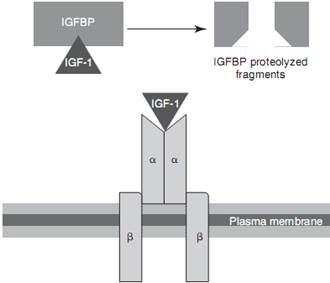
ИФР-1 – простой одноцепочечный полипептид, состоящий из 70 аминокислот, с тремя дисульфидными мостиками и молекулярной массой примерно 7,5 кДа. По структуре он в значительной мере гомологичен проинсулину (Rinderknecht and Humble, 1978a; Cooke et al, 2011; Frystyk, 2012). Структура молекулы ИФР-1, по-видимому, осталась в основном неизменной у разных видов, включая собак и кошек (Zangger et al, 1987; Delafontaine et al 1993). В отличие от инсулина, ИФР-1 связывается группой из шести белков с высоким сродством (ИФР-связывающий белок-1 [ИФРСБ-1] – ИФРСБ-6), и в свободной форме присутствует менее 1% циркулирующего ИФР-1. ИФРСБ-3 – самый важный связывающий белок, который связывает большую часть ИФР-1 (примерно 80-85%) в комплексе с кислотонеустойчивой субъединицей (КНС). Сходный менее распространенный комплекс также образуется с ИФРСБ-5. Эти три молекулы (ИФР-1, ИФРСБ-3/ИФРСБ-5, КНС) образуют крупные трехкомпонентные комплексы массой 150 кДа (Le Roith et al, 2001; Cooke et al 2011). КНС представляет собой гликопротеин массой 85 кДа, образующийся почти исключительно в печени под контролем ГР, откуда он секретируется в кровь. Его основная функция заключается в удлинении времени полувыведения трехкомпонентных комплексов, которые не могут проникнуть через эндотелий сосудов. И наоборот, малое количество ИФР-1 связывается с другими ИФРСБ, образуя двойные комплексы 50 кДа, легко проникающие через эндотелиальный барьер. Время полувыведения свободного ИФР-1 примерно 10 минут, а при образовании тройных комплексов оно увеличивается до более 12 часов. Сообщается, что время полувыведения двойных комплексов составляет 30 минут (Guler et al, 1989a; Boisclair et al, 2001; Cooke et al, 2011). Связывание большего количества ИФР-1 в крупные тройные комплексы позволяет «запасать» высокие концентрации ИФР-1 в циркулирующей крови, предотвращая его инсулиноподобную активность, которая в противном случае привела бы к гипогликемии (Zapf et al 1995). Суммарно, ИФРСБ можно считать белками-носителями ИФР-1 и регуляторами действия ИФР-1 посредством модулирования его доступности. Интересно отметить, что ИФРСБ могут обладать собственной биологической активностью независимо от ИФР-1 (Firth and Baxter, 2002). ИФР-1 диссоциирует, отделяясь от ИФРСБ за счет протеолиза, действия массы или других еще не известных механизмов; в последующем ИФР-1 связывается и активирует рецептор, повсеместно экспрессирующийся на поверхности клеток (рис. 2-3). Рецептор ИФР-1 очень сходен с рецептором инсулина; оба они принадлежат к семейству трансмембранных тирозинкиназ. По структуре они являются тетрамерами, состоящими из двух внеклеточных α-субъединиц с участками связывания лиганда и двух главным образом внутриклеточных β-субъединиц. Последние обладают активностью тирозинкиназы, после активации рецептора инициирующей несколько сигнальных путей (Le Roith et al, 2001; Cooke et al 2011; Frystyk, 2012). Рецептор ИФР-1 связывается с ИФР-1 с высоким сродством; кроме того, он связывает инсулин, но сродство к связыванию в 100 раз ниже. ИФР-1 также может связываться с рецептором инсулина, но с меньшим сродством (Cooke et al, 2011; Bach et al, 2013). Показано, что у собак состав ИФРСБ сходен с таковым у других видов, и ИФРСБ-3 также является самым распространенным связывающим белком (Maxwell et al, 1998). У кошек, по-видимому, формируется меньше тройных комплексов ИФРСБ-3, и предполагают, что КНС является лимитирующим фактором (Lewitt et al, 2000).
|
|
|
|
|
рецептор ИФР-1
РИС. 2-3.Упрощенное схематическое представление диссоциации инсулиноподобного фактора роста-1 (ИФР-1) от ИФР-связывающего белка (ИФРСБ) и связывания свободного ИФР-1 с его рецептором. (Воспроизведено с изменениями из публикации: Cooke DW, et ah Normal and aberrant growth. In Melmed S, Poonsky KS, Larsen PR, Kronenberger HM, editors: Williams'textbook of endocrinology, ed 12, Philadelphia, 2011, Saunders/Elsevier.)
ИФР-2 – одноцепочечный полипептид с молекулярной массой примерно 7,5 кДа, состоящий из 67 аминокислот. Как и ИФР-1, по структуре он сходен с проинсулином и циркулирует в плазме в форме комплексов с шестью ИФРСБ с высоким сродством к связыванию (Frystyk, 2012). Однако рецептор ИФР-2 по структуре не гомологичен рецептору ИФР-1 или инсулина. В то время как рецептор ИФР-1 связывается с обоими ИФР с высоким сродством, рецептор ИФР-2 обладает высоким сродством только к ИФР-2. ИФР-1 обладает меньшим сродством к рецептору ИФР-2, а инсулин не связывается с ним совсем (Cooke et al 2011).
Дата добавления: 2016-09-20; просмотров: 1243;