РЕГУЛЯЦИЯ СЕКРЕЦИИ ГОРМОНА РОСТА В ГИПОФИЗЕ
Синтез и секреция ГР в гипофизе регулируется сложной сетью стимулирующих и подавляющих факторов. Все эти факторы в сочетании приводят к импульсному высвобождению ГР из соматотропных клеток передней доли гипофиза (Mol and Meij, 2008). Таким образом, рилизинг-гормон гормона роста (РГГР) и соматостатин (также называемый фактором, подавляющим высвобождение соматотропина [SRIF]) играют преобладающую роль и относятся к наиболее известным регуляторам. Оба гормона вырабатываются в нейронах гипоталамуса, откуда транспортируются по нервным волокнам во внешнюю пластинку срединного возвышения. При последующем высвобождении в капилляры гипоталамо-гипофизарной портальной системы они достигают передней доли гипофиза (Meij et al 2010a). РГГР – пептид, состоящий из 44 аминокислот и образующийся в результате посттрансляционной модификации более крупной молекулы прогормона. Ее структура несколько варьирует у разных видов. Рецептор РГГР принадлежит к подсемейству рецепторов, связанных с G-белком, и активирует соматотропные клетки посредством активации путей с участием аденилатциклазы/циклического аденозиномонофосфата (цАМФ). Активация рецепторов ведет к высвобождению уже образовавшегося ГР, а также стимуляции генной транскрипции и синтезу нового ГР (Mayo et al, 2000; Low, 2011). РГГР в гипофизе действует почти исключительно на секрецию ГР, и, по-видимому, не взаимодействует с другими рилизинг-гормонами гипоталамуса (Giustina and Veldhuis, 1998; Low, 2011). Время полувыведения РГГР короткое (у человека от 2 до 4 минут) (Javorsky et al 2011).
Соматостатин – мощный ингибитор ГР и снижает как базальную концентрацию, так и частоту и амплитуду импульсного высвобождения под действием РГГР. Он не влияет на синтез ГР. Хотя в этом участвует много других факторов, в настоящее время полагают, что импульсное высвобождение ГР отражает преимущественно импульсную секрецию РГГР, в то время как снижение концентрации ГР контролируется преимущественно соматостатином. Возможно, что высокая концентрация соматостатина в периоды низкой концентрации ГР подготавливает соматотропные клетки к максимальной реакции на последующие импульсные выбросы РГГР (Melmed et al, 2011). Соматостатин обнаруживается не только в гипоталамусе, но и в различных других областях центральной и периферической нервной системы, а также в желудочно-кишечном тракте, поджелудочной железе и других тканях. Следовательно, кроме сильного подавления ГР, он подавляет и другие гормоны (например, ТСГ, инсулин, глюкагон, гастрин), а также иммунные функции и опухолевый рост, и, кроме того, оказывает значимое влияние на функцию желудочно-кишечного тракта и других органов (Reichlin, 1983; Giustina and Veldhuis, 1998). Соматостатин образуется при расщеплении прогормона на тетрадекапептид соматостатин 14 и молекулу соматостатина из 28 аминокислот, называемую соматостатином 28. Соматостатин 14 – основная форма в головном мозге, включая гипоталамус, идентичная у всех позвоночных; соматостатин 28 – основная форма в желудочно-кишечном тракте (Low, 2011). Известно пять рецепторов соматостатина (SSTR1 - SSTR5), все из которых экспрессируются в гипофизе (Giustina and Veldhuis, 1998). Связывание с рецептором соматостатина ведет к активации связанного с мембраной ингибирующего G-белка, подавлению активности аденилатциклазы и снижению внутриклеточной концентрации цАМФ (Low, 2011). Время полувыведения соматостатина примерно так же мало, как у РГГР (Javorsky etal, 2011).
В сложном взаимодействии сигналов обратной связи участвует четыре пептида: ГР, ИФР-1, РГГР и соматостатин (рис. 2-4). ГР сам по себе стимулирует высвобождение соматостатина в гипоталамусе, подавляет РГГР и напрямую подавляет соматотропные клетки. РГГР и соматостатин способны подавлять свою собственную секрецию и взаимно контролировать секрецию своего аналога (Giustina and Veldhuis, 1998). ИФР-1 подавляет секрецию ГР на уровне гипоталамуса, усиливая высвобождение соматостатина, а также непосредственно на уровне гипофиза (Melmed et al, 2011).
За импульсную секрецию ГР ответственны не только РГГР и соматостатин. Сложный ряд внешних и внутренних стимулов (таких как состав тела и упитанность, возраст, пол, сон, физические нагрузки, голодание, генетические основы, заболевания и многие другие) регулирует амплитуду и частоту высвобождения ГР. Значимость этих факторов может различаться в зависимости от вида; например, у собак секреция ГР, по-видимому, не связана со сном или циклами дня и ночи, а индуцированная инсулином гипогликемия не всегда приводит к секреции ГР (French et al, 1987; Bhatti et al, 2006a; Mol and Meij, 2008; рамка 2-1).
Пожалуйста, обратите внимание на вариабельность между видами, например, гипотиреоидизм у собак связан с повышением высвобождения ГР, а секреция ГР, по-видимому, не связана со сном или циклами сна и бодрствования.
модулирующие факторы
| ИФРСБ-1
ИФРСБ-2
ИФРСБ-3
ИФРСБ-4
ИФРСБ-5
ИФРСБ-6
| |
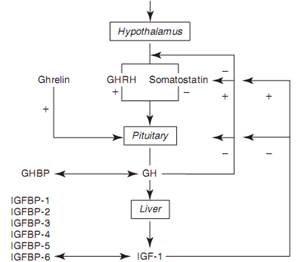
РИС. 2-4.Упрощенное схематическое представление системы гипоталамуса-гипофиза-гормона роста (ГР)/инсулиноподобного фактора роста (ИФР). Подробнее о модулирующих факторах см. в рамке 2-1. БСГР, белок, связывающийся с гормоном роста; РГГР, рилизинг-гормон гормона роста; ИФРСБ, белок, связывающийся с ИФР.
| 2-1. Факторы, модулирующие высвобождение гормона роста у человека
|
| Стимулирующие факторы
| Подавляющие факторы
|
| Физиологические
| Физиологические
|
| Сон
Физические нагрузки
Стресс
Снижение концентрации глюкозы в крови после еды
| Гипергликемия после еды
Повышение концентрации свободных жирных кислот
Старение
|
| Патологические
| Патологические
|
| Истощение запасов белка
Голодание
Хроническая почечная недостаточность
Цирроз печени
Диабет 1 типа
Интерлейкины 1,2,6
| Ожирение
Гипотиреоидизм
Гипертиреоидизм
Гипоадренокортицизм
|
| Фармакологические
| Фармакологические
|
| Гипогликемия под действием инсулина
Инфузия аминокислот (аргинина, лизина)
РГГР
Грелин
АКТГ, α-МСГ, вазопрессин
Эстроген
Агонисты α-адренергических рецепторов (например, клонидин)
Антагонисты β-адренергических рецепторов
Агонисты дофамина
Предшественники серотонина
Агонисты ГАМК
Инфузия калия
| Соматостатин
ИФР-1, ИФР-2
Глюкокортикоиды (длительное введение)
Антагонисты α-адренергических рецепторов
Агонисты β-адренергических рецепторов
Антагонисты серотонина
Антагонисты дофамина
|
Источник: Javorsky BR, et al.: Hypothalamus and pituitary gland. In Gardner DG, Shoback D, editors: Greenspan's basic & clinical endocrinology, ed 9, San Francisco, 2011, McGraw Hill and Low MJ: Neuroendocrinology. In Melmed S, Polonsky KS, Larsen PR, Kronenberger HM, editors: Williams' textbook of endocrinology ed 12, Philadelphia, 2011, Saunders Elsevier, с изменениями.
α-МСГ, α-меланоцитостимулирующий гормон; АКТГ, адренокортикотропный гормон; ГАМК, гамма-аминомасляная кислота; РГГР, рилизинг-гормон ГР; ИФР, инсулиноподобный фактор роста.
Приблизительно 15 лет назад был обнаружен грелин, еще один важный фактор в сложной сети (Kojima et al, 1999). Ghre – индоевропейский корень слова, означающего рост, а «relin» означает высвобождение (Bhatti et al, 2006a). Грелин – пептид длиной 28 аминокислот с высоким структурным сходством между видами. Он циркулирует в крови в неацилированной и ацилированной форме; ацилирование необходимо для биологического действия (Nass et al, 2011). Основным местом синтеза грелина является дно желудка, однако синтез обнаружен также в тонком кишечнике и многих других тканях, включая поджелудочную железу, гипофиз и гипоталамус (Nass et al, 2011; Sato et al, 2012). Грелин действует посредством рецептора, отдельного от рецептора РГГР, называемого рецептором стимулятора секреции ГР (РССГР), или рецептором грелина. Это типичный рецептор, связанный с G-белком, активно экспрессируется в передней доле гипофиза, гипоталамусе, гиппокампе и других областях мозга; его структура осталась в значительной мере неизменной у разных видов позвоночных (Sato et al 2012). Интересно отметить, что РССГР был обнаружен за несколько лет до открытия его естественного лиганда грелина, поэтому назван РССГР-«сиротой». Изначально было обнаружено, что энкефалины обладают свойствами рилизинг-гормона ГР, что стало толчком к разработке синтетических соединений, индуцирующих секрецию ГР (стимуляторы секреции ГР, ССГР). После определения рецептора ССГР проводились активные исследования по поиску его эндогенного лиганда (Koijma et al, 1999; Sato et al 2012).
Грелин действует непосредственно на уровне гипофиза, стимулируя высвобождение ГР (см. рис. 2-4). In vivo эффект сильнее, чем in vitro, что дает основания предполагать синергическое действие грелина и РГГР (т. е. РГГР необходим для индукции максимального высвобождения ГР) (Sato et al, 2012). В силе действия и эффекте других гормонов гипофиза обнаружены межвидовые различия. У людей высвобождение ГР после введения грелина более выражено, чем после введения РГГР, и этот эффект не абсолютно специфичен, поскольку грелин усиливает также секрецию пролактина, АКТГ и кортизола (Bhatti et al, 2006a). У собак высвобождение грелина, по-видимому, зависит от возраста (Yokoyama et al, 2005; Bhatti et al, 2006b). У молодых собак (13-17 мес.) эффект грелина в виде высвобождения ГР сильнее, чем у РГГР, в то время как у собак старшего возраста (7-12 лет) секреция ГР гораздо сильнее стимулировалась РГГР, чем грелином (Bhatti et al, 2006b). У собак грелин не активирует систему гипофиза-коры надпочечников и не стимулирует высвобождение пролактина, ТСГ и ЛГ (Bhatti et al, 2002). У кошек грелин также стимулирует высвобождение ГР в зависимости от дозы (Ida 2012). До настоящего времени сила его действия, зависимость от возраста и специфичность в отношении соматотропных клеток у кошек не исследованы. Грелин обладает различными дополнительными физиологическими функциями, такими как регуляция аппетита (индукция голода), стимуляция моторики желудка и секреции кислоты, снижение секреции инсулина, а также влияет на сердечно-сосудистую систему и оказывает антипролиферативное действие (Sato et al, 2012).
И наконец, синтез и высвобождение ГР модулируется другими гормонами, например гормонами щитовидной железы, глюкокортикоидами и половыми гормонами. У людей первичный гипотиреоидизм сопровождается ослаблением секреции ГР. Однако у собак с гипотиреозом концентрации базального ГР и ИФР-1 повышены (Lee et al, 2001). У собак с гипофизарным гиперадренокортицизмом базальные концентрации ГР нормальны, однако частота импульсов высвобождения и количество секретируемого ГР меньше. У людей также отмечено ингибирующее действие длительного избытка глюкокортикоидов, возможно, частично опосредованное усилением высвобождения соматостатина (Lee et al 2003).
Дата добавления: 2016-09-20; просмотров: 1319;