МЕТАБОЛИЧЕСКОЕ ДЕЙСТВИЕ ГОРМОНА РОСТА И ИНСУЛИНОПОДОБНОГО ФАКТОРА РОСТА-1
Действие ГР и ИФР-1 обычно описывают как отдельные явления (т. е. быстрое прямое действие приписывают ГР, а медленное косвенное - ИФР-1). Однако представления эволюционировали, и в настоящее время установлено, что между двумя гормонами существуют тесные и гораздо более сложные взаимодействия. Несколько исследований показало взаимное влияние между путями, активируемыми ГР, ИФР-1 и инсулином (Vijayakumar et al, 2010). В общих чертах, ГР оказывает свое действие посредством связывания с рецептором ГР, который обильно экспрессируется в печени, а также в других тканях. Многие его эффекты опосредованы последующей выработкой ИФР-1 (рис. 2-5). Печень – преимущественный источник ИФР-1, хотя он синтезируется во многих тканях, где действует аутокринным/паракринным способом (Aragon-Alonso et al, 2011). ГР стимулирует выделение ИФР-1 в циркулирующую кровь при достаточном поступлении питательных веществ и высокой концентрации инсулина в воротной вене, в то время как во время голодания концентрация ИФР-1 снижается (Moller and Jorgensen, 2009). У взрослых ГР является основным метаболическим гормоном, оптимизирующим состав тела и физическую функцию, а также регулирующим обмен веществ и энергии. Он тесно взаимодействует с инсулином при регуляции обмена жиров, белков и углеводов во время голодания и после приема пищи (Melmed et al, 2011). ГР стимулирует расщепление жиров и окисление жирных кислот, что особенно важно при голодании для получения энергии без расщепления белков. Последний эффект (способствующий сохранению белка) – важный механизм, по которому ГР способствует росту и развитию (Melmed et al, 2011; Javorsky et al 2011). ГР также играет роль в дифференциации жировых клеток; появление крупных зрелых жировых клеток связано с повышенной способностью к запасанию триглицеридов и большей липофильной активностью (Vijayakumar et al, 2010). ГР усиливает синтез белка (непосредственно и посредством ИФР-1), усиливая все механизмы усвоения аминокислот и их встраивания в белки при одновременном подавлении расщепления белков. Влияние ГР на обмен углеводов как прямое, так и косвенное. Прямое действие ГР, противоположное эффекту инсулина, особенно важно во время голодания и заключается в снижении усвоения глюкозы в тканях за пределами печени, усилении образования глюкозы в печени и повышении концентрации глюкозы в плазме (Sjaastad et al, 2010; Javorsky et al, 2011). ИФР-1 влияет на углеводный обмен очень разными путями. Его эффекты подобны эффектам инсулина и включают усиление усвоения глюкозы в тканях за пределами печени, снижение выработки глюкозы в печени и снижение концентрации глюкозы в плазме (Mauras and Haymond, 2005; Javorsky et al, 2011). В случае избытка ГР антиинсулиновое «диабетогенное» действие может привести к клиническому диабету и возможной резистентности к инсулину.
ГР

| Антагонист инсулина | Расщепле-ние жиров | Синтез белка | Рост эпифизов |
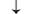
ИФР-1

| Инсулино-подобное действие | Антилипо-литическое действие | Синтез белка | Рост эпифизов |
РИС.2-5 Упрощенная схема основного метаболического действия гормона роста (ГР) и инсулиноподобного фактора роста (ИФР-1).
Растущим индивидам необходима нормальная секреция ГР и функциональная целостность системы ГР-ИФР-1 для нормального роста в длину и развития скелета. В медицине соответствующие роли ГР, ИФР-1, их связывающих белков и рецепторов, а также влияние других факторов (например, цитокинов) интенсивно исследуются, и на данный момент описано впечатляющее количество генетических дефектов (Savage et al 2011). Полагают, что ГР действует на предшественников зоны роста, стимулируя дифференциацию хондроцитов и локальный синтез ИФР-1, что, в свою очередь, действует аутокринным/парактинным способом и индуцирует клональную пролиферацию и гипертрофию хондроцитов. Кроме того, возможно, что ГР оказывает дополнительное прямое действие на пролиферацию и гипертрофию хондроцитов (Zezulak and Green, 1986; Gevers et al, 2002). Эффект усиления роста также частично опосредуется циркулирующим ИФР-1, образовавшимся в печени (Stratikopoulos et al, 2008).
Собаки – уникальный вид млекопитающих, так как разница в массе животных, находящихся на противоположных концах диапазона (например, чихуахуа и сенбернара), достигает почти 100 раз (Favier et al, 2001; Rijn-berk et al, 2003). Начальные исследования показали различные концентрации ИФР-1 в циркулирующей крови взрослых собак разных пород; у мелких пород концентрации ИФР-1 ниже, чем у средних, у которых они, в свою очередь, ниже, чем у крупных (Eigenmann et al, 1984a). Сходным образом, в генетических подгруппах в пределах одной породы концентрации ИФР-1 соответствуют размеру, так как у стандартных пуделей концентрации ИФР-1 гораздо выше, чем у карликовых пуделей, в то время как концентрации ГР сходны (Eigenmann et al, 1984c). Другое исследование большого числа собак многих пород подтвердило зависимость между ИФР-1 и размером тела, поскольку концентрация ИФР-1 возрастала с увеличением массы (Greer et al, 2011). Однако результаты оценки молодых растущих собак заставили усомниться в представлении, что концентрация ИФР-1 в циркулирующей крови является основной определяющей размера собак (Rijnberk et al, 2003). Длительное введение щенкам карликового пуделя рекомбинатного человеческого ИФР-1 не привело к увеличению размеров по сравнению с контрольными собаками несмотря на значительное повышение концентрации ИФР-1 в циркулирующей крови (Guler et al, 1989b). Между концентрацией ИФР-1 и скоростью роста щенков стандартного и карликового пуделя не обнаружено корреляции (Biirki, 2000).
Вопрос связи ИФР-1 и размера тела дополнительно изучался при генетических исследованиях. Показано, что ген ИФР-1, как ген-кандидат, обусловливающий размер тела, вносит основной вклад в разнообразие размеров у собак. Ген ИФР-1 и связанные гены отвечают примерно за 50% генетической вариабельности размеров собак (Sutter et al, 2007; Boyko, 2011; Wayne and von Holdt, 2012). Мелкие породы собак имеют гаплотипы ИФР-1, близкородственные таковым у волков Среднего Востока, что дает основания предполагать раннюю эволюцию собак малого размера в этом регионе (Gray et al, 2010; Wayne and vonHoldt, 2012). Очевидна роль дополнительных факторов; вероятно, что большую роль играет ГР. Показано, что у щенков дога базальные концентрации, а также импульсы высвобождения ГР значительно выше, чем у биглей такого же возраста. У биглей концентрация ГР снизилась до диапазона, характерного для взрослых собак, уже к 7 неделе, в то время как у догов снижение происходило гораздо медленнее: концентрации ГР оставались повышенными на протяжении 24 недель. Следовательно, авторы предположили, что разная длительность сохранения повышенной концентрации ГР может отвечать за различия в массе тела у собак средних и крупных пород (Favier et al, 2001).
Дата добавления: 2016-09-20; просмотров: 750;