БИОСИНТЕЗ ГОРМОНА РОСТА И ИНСУЛИНОПОДОБНОГО ФАКТОРА РОСТА-1
Нарушения гормона роста
РАЗВИТИЕ ГИПОФИЗА
Гипофиз играет центральную роль в регуляции роста. Он состоит из двух основных частей, аденогипофиза и нейрогипофиза, имеющих разное эмбриональное происхождение. Аденогипофиз, также называемый передней долей гипофиза, состоит из дистальной, промежуточной и воронковидной частей. Структура, дающая начало аденогипофизу, была впервые описана немецким анатомом, эмбриологом и зоологом Ратке в 1838 году и названа в его честь. В период эмбриогенеза аденогипофиз развивается из кармана Ратке, представляющего собой выпячивание слоя эктодермы толщиной в одну клетку на крыше примитивной ротовой полости. Этот слой мигрирует, чтобы присоединиться к нейроэктодерме зачатка вентральной части гипоталамуса; контакт важен, так как индуцирующие сигналы от гипоталамуса необходимы для нормального развития передней доли гипофиза. В последующем карман Ратке структурно отделяется от ротовой полости, передняя стенка утолщается и формирует дистальную часть аденогипофиза. Задняя стенка кармана Ратке формирует промежуточную часть, которая остается отделенной от дистальной части гипофизарной щелью. Нейрогипофиз происходит из нервной эктодермы и представляет собой продолжение вентральной части гипоталамуса. У собак и кошек аденогипофиз образует ободок вокруг проксимальной части нейрогипофиза, а также окружает часть срединного возвышения (Meij, 1997; Kooistra, 2000; Meij etal, 2010a; рис. 2-1).
Развитие гипофиза и детерминация специфических клеточных типов из общих клеток-предшественников происходят в определенном и четко регулируемом порядке. Эти процессы контролируются сложным каскадом факторов транскрипции и сигнальных молекул. После пролиферации появляются разные типы клеток с соответствующим пространственным и временным распределением, подвергающиеся высокоселективной дифференциации. Кортикотропные клетки – первый определенный клеточный тип в созревающей передней доле гипофиза (Kooistra et al, 2000a; Kelberman et al, 2009; Javorsky et al 2011; рис. 2-2). У людей изменения этих процессов и путей связаны с большим спектром заболеваний и являются темой и областью интенсивных исследований; у собак и кошек изучение роли возможных мутаций началось только недавно. Клетки передней доли изначально классифицировались по их реакции с разными красителями как ацидофилы, базофилы и хромофобы. В настоящее время иммунологические техники мечения позволяют нам классифицировать клетки гипофиза по специфическим продуктам их секреции (Childs, 2009). Различают пять типов эндокринных клеток: кортикотропные клетки (секретирующие адренокортикотропный гормон [АКТГ] и родственные пептиды), тиреотропные (секретирующие тиреостимулирующий гормон [ТСГ]), гонадотропные клетки (секретирующие лютеинизирующий гормон [ЛГ] и фолликулостимулирующий гормон [ФСГ]), соматотропные клетки (секретирующие гормон роста [ГР]) и лактотропные клетки (секретирующие пролактин). Соматотропные и лактотропные клетки ацидофильные, а тиреотропные и гонадотропные – базофильные. К хромофобным клеткам относятся кортикотропные, несекретирующие фолликулярные (звездчатые) клетки, дегранулированные хромофильные и недифференцированные стволовые клетки (Capen, 2007).
|
|
|
|
|
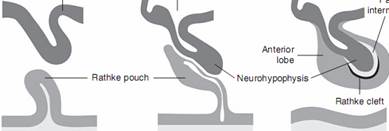
РИС. 2-1.Упрощенная иллюстрация онтогенеза гипофиза. (Воспроизведено с разрешения из публикации: Meij BP, et ah Hypothalamus-pituitary system. In Rijnberk A, Kooistra HS, editors: Clinical endocrinology of dogs and cats, ed 2, Hannover, 2010, Schliitersche)
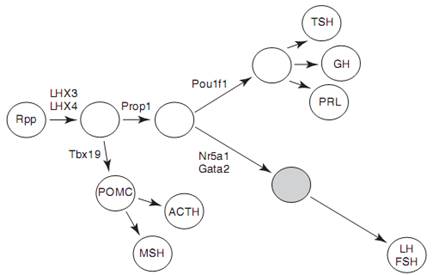
РИС. 2-2.Упрощенное, схематическое представление дифференциации клеточных линий гипофиза. Окончательные типы эндокринных клеток помечены синтезирующимися в них гормонами. В процессе клеточной дифференциации участвуют различные факторы транскрипции. Показаны некоторые из наиболее известных факторов. (Воспроизведено с изменениями из публикаций: Genetic regulation of pituitary gland development in human and mouse, Endocr Rev 30[7]:790-829, 2009; и Metherell LA, et al Genetic defects of the human somatotropic axis. In Wass JAH, Stewart PM, Amiel SA, Davies MJ, editors: Oxford textbook of endocrinology and diabetes, ed 2, Oxford, 2011, Oxford University Press.)
ACTH, адренокортикотропный гормон; FSH, фоллкулостимулирующий гормон; Gata2, gata-связывающий белок 2; GH, гормон роста; LH, лютеинизирующий гормон; LHX3/LHX4, фактор транскрипцииLIM-домена; MSH, меланоцитостимулирующий гормон; Nr5al, ядерный рецептор подсемейтва 5, группа A, член 1 (также называемый стероидогенным фактором-1 [SF1]); POMC, проопимеланокортин; Poulfl (также называемый PIT1) гипофизарный положительный фактор транскрипции 1; PRL, пролактин; Propl, фактор транскрипции Prophet PIT1; Rpp, предшественник кармана Ратке; Tbxl9; фактор транскрипции T-box 19 (ранее называвшийся гипофизарным фактором транскрипции T-box (TPIT)), TSH, тиреостимулирующий гормон.
Соматотропные клетки – наиболее обильный тип, составляющий примерно 50% клеток передней доли. Процентная доля других клеточных типов варьирует от 10 до 15%. Распределение клеточных типов не случайно, а обусловливается топологической и количественной организацией. Соматотропные клетки расположены главным образом в дорсальной области дистальной части, близко к промежуточной части (Meij et al, 2010b). Однако за последние два десятилетия стало очевидно, что традиционная концепция, предполагающая, что каждый тип клеток гипофиза запасает и секретирует только один гормон и регулируется специфическим рилизинг-фактором гипоталамуса, слишком узка и существуют дополнительные механизмы (Childs, 2009; Meij et al, 2010b). В пределах одного типа эндокринных клеток существуют субпопубяции, синтезирующие несколько гормонов и способные активироваться несколькими рилизинг-гормонами. Примерами являются маммосоматотропные клетки, способные запасать и высвобождать ГР и пролактин, соматогонадотропные клетки, способные запасать и высвобождать ГР и ЛГ/ФСГ, тиреосоматотропные клетки, содержащие ТСГ и ГР. При этом возможно фенотипическое «переключение» между двумя соответствующими типами зрелых клеток в зависимости от потребности организма; это явление называется трансдифференциацией (Kineman et al, 1992; Childs, 2000; Vidal et al, 2000; Villalo-bos et al, 2004; Childs, 2009). Большинство исследований проводилось на экспериментальных животных, культурах клеток или биоптатах, взятых у людей, поэтому знания о возможной трансдифференциации клеток гипофиза у собак и кошек скудны. Тем не менее, недавнее исследование дает основания предполагать существование сходных механизмов. За группой собак породы бигль наблюдали в течение 3 лет после индукции первичного гипотиреоидизма. За это время базальная концентрация ГР повысилась, а при стимуляции тиреотропин-рилизинг-гормоном было отмечено парадоксальное усиление реакции. Результаты гистологического и иммуногистохимического исследования гипофиза показали гиперплазию тиреотропных клеток, крупные вакуолизированные «клетки недостаточности щитовидной железы» и клетки гипофиза, окрашивающиеся положительно как на ГР, так и на ТСГ; последнее указывает на трансдифференциацию соматотропных клеток в тиреотропные (Diaz-Espifieira et al, 2008).
БИОСИНТЕЗ ГОРМОНА РОСТА И ИНСУЛИНОПОДОБНОГО ФАКТОРА РОСТА-1
Дата добавления: 2016-09-20; просмотров: 1209;