КРАТКИЙ ОЧЕРК ИСТОРИИ АНАТОМИИ 60 страница
ЕДИНСТВО ВЕГЕТАТИВНОЙ И АНИМАЛЬНОЙ
ЧАСТЕЙ НЕРВНОЙ СИСТЕМЫ
Необходимо помнить, что вегетативная нервная система есть часть единой нервной системы. Поэтому в целом организме постоянно наблюдается сочетанная деятельность вегетативной и анимальной частей нервной системы с вовлечением центров, находящихся на различных уровнях нервной системы.
Рассмотрим такую сочетанную деятельность на примере регуляции акта мочеиспускания.
В едином акте мочеиспускания участвуют непроизвольные мышцы (m. detrusor vesicae и т. sphincter vesicae), иннервируемые вегетативными нервами, и произвольная (m. sphincter urethrae), иннервируемая анимальными. При этом вытеснитель мочи сокращается, а оба сфинктера расслабляются, хотя каждый из них иннервируется из разных частей нервной системы:
|
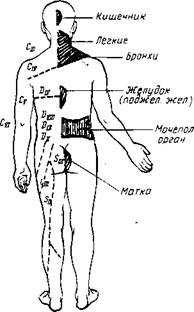
| Рис. 347. Зоны отраженных болей (зоны Захарьина — Геда) при заболеваниях внутренних органов. |



 сфинктер пузыря — из вегетативной, а сфинктер мочеиспускательного канала — из анимальной. Это происходит благодаря наличию общего центра координации в головном мозге, поскольку вегетативная и анимальная части составляют единую нервную систему.
сфинктер пузыря — из вегетативной, а сфинктер мочеиспускательного канала — из анимальной. Это происходит благодаря наличию общего центра координации в головном мозге, поскольку вегетативная и анимальная части составляют единую нервную систему.
В интеграции анимальной и вегетативной нервных систем большую роль играет лимбическая система.
Зоны Захарьина —Геда. В настоящее время имеются сведения об афферентной иннервации внутренностей анимальной нервной системой, что отмечалось выше. Возможно, этим объясняется давно известный симптом отраженных болей, наблюдаемый в клинике. Заболевания некоторых внутренних органов постоянно сопровождаются отраженными болями в определенных местах кожи. Таковы, например, боли в левой лопатке и левой руке при стенокардии, боли между лопатками при язве желудка, боли в правой подвздошной ямке при аппендиците и др. Эти боли локализуются в определенных кожных сегментах, соответствующих тем сегментам спинного мозга, куда поступают афферентные (чувствительные) волокна из пораженного внутреннего органа. Такие кожные сегменты, или зоны, называются зонами Захарьина — Геда по имени, описавших их авторов. Приводим схему этих зон (рис. 347). Знание зон Захарьина — Геда помогает судить по болям в наружных покровах тела о состоянии органов внутри его полостей. Возможно, что зоны Захарьина — Геда обусловливают воздействие на внутренние органы применяемых в китайской медицине прижиганий или вкалываний игл (акупунктура) в определенные кожные точки.
Сегментарная иннервация органов. Внутриорганные нервы паренхиматозных органов распределяются, как и сосуды, соответственно сегментам органов. Так, в легких различают 10 нервных сегментов. В печени число их весьма варьирует и может достигать восьми. В почке отмечается 5 сегментов иннервации.
ОБЩИЙ ОБЗОР ОСНОВНЫХ ПРОВОДЯЩИХ ПУТЕЙ НЕРВНОЙ СИСТЕМЫ
Как уже отмечалось, интеграция организма в единое целое осуществляется нейрогуморальной регуляцией при ведущей роли нервной системы. Нервная система обеспечивает и единство организма и среды. Рассмотрим морфологическую основу этой интеграции.
В основе деятельности нервной системы лежит рефлекторная дуга. Короткая рефлекторная дуга (см. рис. 264) построена следующим образом. На первом этапе развития центральной нервной системы, когда не было еще головного мозга, рефлекторная дуга замыкалась только в пределах туловищного мозга. Как отражение этого этапа, у человека сохранился собственный аппарат спинного мозга, построенный по принципу трехчленной рефлекторной дуги.
Первый (афферентный, чувствительный) нейрон этой дуги представлен клетками спинномозгового узла, периферические отростки которых идут в составе нервов от органов и тканей, где начинаются рецепторами, а центральные входят в составе задних корешков в спинной мозг. Каждый центральный отросток, войдя в белое вещество спинного мозга, Т-образно разделяется на две ветви — восходящую и нисходящую, от которых в свою очередь отходит несколько боковых веточек (коллатералей). Все эти веточки заканчиваются в задних рогах и substantia intermedia centralis серого вещества нескольких соседних сегментов. Лежащие здесь клетки являются вторым (замыкательным) нейроном простой рефлекторной дуги. Отростки их также делятся на восходящую и нисходящую ветви с боковыми коллатералями, заканчивающимися на клетках передних рогов в пределах нескольких соседних сегментов. Клетки передних рогов составляют третий (эфферентный, двигательный) нейрон; отросток его выходит из спинного мозга в составе передних корешков и далее в составе нервов достигает эффекторов. В результате такого строения простой рефлекторной дуги один чувствительный нейрон вступает в связь с несколькими промежуточными нейронами, а через их разветвления — с еще большим числом двигательных нейронов, вследствие чего раздражение из одной точки тела может передаваться не только на соответственный сегмент, но и на ряд ближайших. Благодаря этому простой рефлекс может стать более распространенным, с вовлечением в ответную реакцию большой группы мышц.
В спинном мозге человека имеются и двучленные дуги, лишенные промежуточного нейрона (см. рис. 351). Примером может служить сухожильный коленный рефлекс, вызываемый постукиванием молоточка по ligamentum patellae при согнутой в колене ноге. В этом случае раздражение передается с сухожильного рецептора на периферический отросток лежащего в спинномозговом ганглии чувствительного нейрона, центральный отросток которого, вступив в составе задних корешков в спинной мозг, достигает серого вещества переднего рога, где и заканчивается на его клетках. Последние образуют второй, двигательный, нейрон, отросток которого в составе переднего корешка и далее мышечного нерва доходит до заложенного в мышцах эффектора. В результате в ответ на постукивание молоточком по сухожилию четырехглавой мышцы бедра наблюдаются ее сокращение и рефлекторное разгибание голени. Двучленная дуга считается молодым приобретением животных, а трехчленный рефлекторный аппарат спинного мозга является филогенетически древним аппаратом. На его базе с возникновением головного мозга стал развиваться более молодой проводниковый аппарат, связывающий спинной мозг с развивающимся головным.
По мере развития головного мозга возникают и разрастаются двусторонние связи спинного мозга с головным, вследствие чего с возникновением каждого нового этажа последнего увеличивается число связанных с ним афферентных и эфферентных нейронов. Рефлекторная дуга усложняется, так что вместо одного нейрона в каждой ее части появляются цепи нейронбв, образующих афферентные и эфферентные проводящие пути. Следовательно, проводящими путями в нервной системе называются тесно расположенные одно возле другого нервные волокна, соединяющие различные отделы ее и объединенные в системы пучков, характеризующиеся общностью строения и функции. С помощью проводящих путей и достигаются единство организма и его связь со средой. Чтобы понять их строение, нужно учитывать основные этапы эволюции центральной нервной системы, последовательное развитие отделов головного мозга (см. «Филогенез»), У человека существуют одновременно и старые, и новые проводящие пути, благодаря которым спинной мозг оказывается связанным со всеми отделами головного мозга (см. рис. 270).
1. С продолговатым мозгом:
а) восходящие — fasciculus gracilis и fasciculus cuneatus, идущие от спинномозговых ганглиев через задние канатики спинного мозга до соименных ядер продолговатого мозга, nucleus gracilis и nucleus cuneatus;
б) нисходящие — от ядер, имеющих отношение к равновесию и координации движений, до передних рогов спинного мозга — tractus vestibulospinalis, tractus reticulospinalis и tractus olivospinalis.
2. С мозжечком:
а) восходящие — tractus spinocerebellaris posterior и tractus spinocerebellaris anterior; они заканчиваются в коре древней части мозжечка, т. е. в черве. Из них задний состоит из отростков клеток nucleus thoracicus задних рогов на своей стороне и входит в мозжечок в составе нижних его ножек — прямой спинно-мозжечковый путь. Передний состоит из отростков клеток substantia intermedia centralis своей и противоположной стороны. Он поднимается до среднего мозга и входит в мозжечок в составе его верхних ножек. Его волокна переходят на противоположную сторону, образуя перекрещенный спинно-мозжечковый путь; б) нисходящие — от мозжечка к спинному мозгу через средний мост и продолговатый мозг.
3. Со средним мозгом:
а) восходящие — tractus spinotectalis, идущий от задних рогов через ствол мозга до крыши (tectum) среднего мозга; на пути он перекрещивается в commissura alba спинного мозга; б) нисходящие (к передним рогам): tractus tectospinalis — от tectum среднего мозга и tractus rubrospinalis — от красного ядра.
4. С передним мозгом:
а) восходящие идут от клеток nucleus proprius задних рогов спинного мозга к таламусу — tractus spinothalamicus lateralis et anterior и перекрещиваются на своем пути в commissura alba спинного мозга; б) нисходящие — tractus thalamospinalis — от таламуса к передним рогам спинного мозга.
С передним мозгом связаны и пути, идущие от нижележащих отделов головного мозга: lemniscus medialis — идет от ядер fasciculus gracilis et cuneatus продолговатого мозга к таламусу, перекрещиваясь на пути в decussatio lemniscorum.
5. С развитием коры большого мозга возникают ее связи с нижележащими отделами, над которыми она становится надстройкой. У чело
века имеются: а) восходящие — tractus thalamocorticalis — от таламуса к коре большого мозга; б) нисходящие — tractus pyramidalis — от коры большого мозга к ядрам черепных нервов, заложенным в мозговом стволе — tractus corticonuclearis, и к передним рогам спинного мозга — tractus corticospinalis (pyramidalis). Чем более развита кора большого мозга, тем более развиты и исходящие из нее пирамидные пути, достигающие наивысшего развития у человека соответственно наивысшему развитию у него новой коры.
Кроме названных основных пучков нервных волокон, возникают и другие пути, связывающие отдельные части спинного и головного мозга между собой. Благодаря им устанавливается единство всей нервной системы.
СХЕМА ПРОВОДЯЩИХ ПУТЕЙ НЕРВНОЙ СИСТЕМЫ
Проводящие пути с точки зрения направления проведения импульса могут быть разделены на две большие группы — афферентные и эфферентные. Афферентные проводящие пути составляют среднее звено — кондуктор того или иного анализатора; поэтому часть их будет рассмотрена вместе с соответствующими анализаторами (см. «Органы чувств»).
АФФЕРЕНТНЫЕ (ВОСХОДЯЩИЕ) ПРОВОДЯЩИЕ ПУТИ
Поскольку организм получает раздражение как из внешней среды, так и из внутренней, имеются пути, несущие импульсы от рецепторов, воспринимающих внешние раздражения, и от рецепторов, воспринимающих внутренние раздражения.
Проводящие пути от рецепторов внешних раздражений
Рецепторы, воспринимающие внешние раздражения, называются экстеро- цепторами. На ранних стадиях эволюции они были заложены главным образом в наружных покровах тела, что необходимо для восприятия внешних раздражений, почему и у человека они развиваются в эмбриогенезе из наружного зародышевого листка — эктодермы. Исключение представляет орган вкуса, тесно связанный функционально с пищеварительной системой и поэтому развивающийся из энтодермы (эпителия глоточных карманов). В дальнейшем с усложнением организации животных и усложнением их образа жизни те из экстероцепторов, которые имели жизненно важное значение, начали усиленно развиваться и усложняться в своей организации, приобретая строение особых органов, воспринимающих раздражения, источники которых находятся на известном расстоянии от организма и потому называемые дистантными. Это — рецепторы слуха, зрения и обоняния. Остальные рецепторы наружных покровов остались заложенными в коже, составляя периферическую часть кожного анализатора. Проводящие пути от рецепторов звука, света, вкуса и обоняния будут рассмотрены при описании соответствующих анализаторов в разделе эстезиологии. Здесь будут изложены проводящие пути кожного анализатора.
Проводящие пути кожного анализатора
Афферентные волокна кожного анализатора несут в кору большого мозга тактильные раздражения, чувство стереогноза (узнавание предмета на ощупь), болевые и температурные раздражения. В связи с этим их можно разбить на несколько групп.
Проводящие пути тактильной чувствительности кожи (чувство осязания)
(рис. 348). Tractus gangliospinothalamocorticalis. Рецептор находится в толще кожи. Кондуктор состоит из 3 нейронов. Клеточное тело первого нейрона помещается в спинномозговом узле, который представляет собой скопление клеток периферических нейронов всех видов чувствительности. Отходящий от клеток этого узла отросток делится на две ветви, из которых периферическая идет в составе кожного нерва от рецептора., а центральная в составе заднего корешка идет в задние канатики спинного мозга, где в свою очередь делится на восходящую и нисходящую ветви-. Концевые разветвления и коллатерали одной части волокон заканчиваются в задних рогах спинного мозга в substantia gelatinosa (эта часть тракта носит название tractus gangliospinalis), другая часть восходящих волокон не заходит в задние porav а идет в задних канатиках спинного мозга и достигает в составе fasciculus gracilis et cuneatus соименных ядер продолговатого мозга, nucleus gracilis и nucleus cuneatus (эта часть тракта называется tractus gangliobulbaris).
В задних рогах спинного мозга и в названных ядрах продолговатого мозга помещается клеточное тело второго нейрона. Аксоны клеток, заложенных в задних рогах, пересекают срединную плоскость в commissura alba и входят в состав расположенного в боковом канатике противоположной стороны tractus spinothalamicus lateralis, который они и образуют (см. рис. 270). '
Важно иметь в виду, что перекрест волокон спинно-таламических пучков происходит не на уровне вступления соответствующего заднего корешка в спинной мозг, а на 2 — 3 сегмента выше. Этот факт имеет существенное значение для клиники, так как при ..одностороннем повреждении этого пучка расстройство кожной чувствительности на противоположной стороне наблюдается не на уровне поражения,'а книзу от него.
Этот пучок через стволовую часть головного мозга достигает таламуса. По пути он устанавливает связь с двигательными ядрами мозгового ствола и черепных нервов, по которым возникают головные рефлексы при раздражении кожи, например движение глаз при раздражении кожи руки. Аксоны клеток второго звена, заложенных в ядрах продолговатого мозга, также достигают таламуса по-тракту, носящему название lemniscus medialis, который в продолговатом мозге переходит на противоположную сторону, 'образуя перекрест медиальной петли (decussatio lemniscorum) (рис. 349). Таким образом, для каждой половины тела в спинном мозге имеются как бы два тракта, передающих импульсы прикосновения: 1) один, неперекрещенный, — в заднем канатике той же стороны и 2) другой, перекрещенный, — в боковом канатике противоположной стороны. Поэтому при одностороннем поражении спинного мозга тактильная чувствительность может оставаться ненарушенной, так как сохраняется соответствующий пучок на здоровой стороне.
В таламусе находится клеточное тело третьего нейрона, аксоны которого направляются в кору большого мозга в составе tr. thalamocor- ticalis, в постцентральную извилину (поля 1, 2, 3) и верхнюю теменную дольку (поля 5, 7), где находится корковый конец кожного анализатора (рис. 350, см. рис. 299).
Тактильная и болевая чувствительность имеет разлитую локализацию в коре головного мозга, что объясняет их меньшее нарушение при ограниченных корковых очагах повреждения.
Проводящие пути пространственной кожной чувствительности—стереогноза (узнавание предметов на ощупь) (см. рис. 270). Этот вид кожной чувствительности имеет, как и тактильная чувствительность, идущая по fasciculus gracilis et cuneatus, три звена: 1) спинномозговые ганглии, 2) пй-
Рис. 349. Проекция хода медиальной петли на латеральную поверхность 4 мозгового ствола.
 lm — lemniscus medialis; I, 2, 3 — поперечные сечения продолговатого мозга, моста и среднего мозга с обозначением положения медиальной петли (/т) в толще этих образований;
lm — lemniscus medialis; I, 2, 3 — поперечные сечения продолговатого мозга, моста и среднего мозга с обозначением положения медиальной петли (/т) в толще этих образований;
4 — nucl. lateralis thalami; 5 — пути заднего канатика спинного мозга; 6 — tr. gangliospi- nothalamicus; 7 — decussatio lemniscorum.
cleus gracilis et cuneatus в продолговатом мозге, 3) таламус и, наконец, корковый конец кожного анализатора в верхней теменной дольке (поля 5, 7).
Проводящие пути болевой и температурной чувствительности.
Клеточное тело первого нейрона лежит в спинномозговом узле, клетки которого связаны периферическими отростками с кожей, а центральными — с задними рогами спинного мозга (nuclei proprii), где помещается клеточное тело второго нейрона (tractus gangliospinalis). Аксон второго нейрона переходит на другую сторону в составе commissura alba и поднимается в составе tractus spinothalamicus lateralis до таламуса. Следует отметить, что tractus spinothalamicus lateralis в свою очередь делится на две части — переднюю и заднюю, из которых по передней передается болевая чувствительность, а по задней — термическая. В таламусе лежит клеточное тело третьего нейрона, отросток которого в составе tractus thalamocorti-
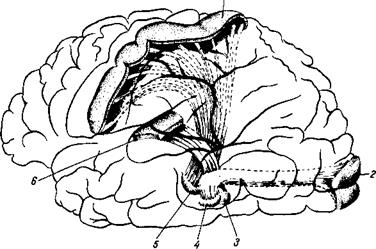 Рис. 350. Схема-модель трех главных чувствительных систем левого полушария большого мозга, восходящих к клеткам его коры.
1 — область общей чувствительности коры полушария; 2 — зрительная область коры полушария; 3 — медиальное коленчатое тело; 4 — латеральное коленчатое тело; 5 — латеральное ядро таламуса; б — слуховая область коры головного мозга.
Рис. 350. Схема-модель трех главных чувствительных систем левого полушария большого мозга, восходящих к клеткам его коры.
1 — область общей чувствительности коры полушария; 2 — зрительная область коры полушария; 3 — медиальное коленчатое тело; 4 — латеральное коленчатое тело; 5 — латеральное ядро таламуса; б — слуховая область коры головного мозга.
|
calis направляется в кору большого мозга, где заканчивается в постцентральной извилине (корковый конец кожного анализатора).
Некоторые полагают, что чувство боли воспринимается не только в коре, но и в таламусе, где различные виды чувствительности приобретают эмоциональную окраску. Болевые и температурные импульсы от отделов или органов головы приходят по соответствующим черепным нервам — V, VII, IX, X парам к их чувствительным ядрам и от этих ядер к таламусу и далее в кору постцентральной извилины (нижний отдел).
Вследствие перекреста волокон второго нейрона проводящих путей, идущих от экстероцепторов, импульсы болевой, температурной и частично тактильной чувствительности передаются в постцентральную извилину с противоположной стороны тела. Поэтому следует запомнить, что поражение первого нейрона или второго нейрона до перекреста вызывает расстройство чувствительности на стороне поражения. Если же пострадали волокна второго нейрона после перекреста или третий нейрон, то расстройство тех же видов чувствительности наблюдается на стороне, противоположной очагу поражения.
Проводящие пути от рецепторов внутренних раздражений
Проводящие пути от рецепторов внутренних раздражений могут быть разделены на проводящие пути от аппарата движения (собственно тела), т. е. от проприоцепторов (proprius — собственный), составляющих кондуктор двигательного анализатора, и пути от рецепторов внутренностей и сосудов, т. е. интероцепторов; вторая группа путей является кондуктором интероцеп- тивного анализатора.
Проводящие пути двигательного анализатора
Двигательный анализатор воспринимает глубокую проприоцептивную чувствительность, к которой относятся мышечно-суставное чувство, вибрационная чувствительность, чувство давления и веса (гравитация). Основной вид проприоцептивной чувствительности — это мышечно-суставное чувство, т. е. импульсы, которые возникают в связи с изменениями степени натяжения суставной сумки, сухожилий и напряжения мышц; благодаря этим импульсам у человека создается представление о положении тела и частей его в пространстве и об изменении этого положения (что имеет, в частности, значение при полетах в космос, где создается состояние невесомости).
Проводящими путями двигательного анализатора являются tractus gangliobulbothalamocorticalis и tractus spinocerebellaris anterior et posterior.
Tractus gangliobulbothalamocorticalis (см. рис. 349). Рецепторы находятся в костях, мышцах, сухожилиях, суставах, т. е. в собственно теле, отчего называются проприоцепторами (рис. 351).
Кондуктор состоит из трех нейронов. Клеточное тело первого нейрона помещается в спинномозговом узле. Аксон этой клетки делится на две ветви — периферическую, идущую в составе мышечного нерва от проприо- цептора, и центральную, идущую в составе задних корешков в задние канатики спинного мозга, fasciculus gracilis и fasciculus cuneatus, до продолговатого мозга (см. рис. 270, 348, 349). Здесь они оканчиваются в соименных ядрах названных канатиков — nucleus gracilis и niicleus cuneatus (tractus gangliobulbaris).
В этих ядрах помещаются тела вторых нейронов. Аксоны их в составе lemniscus medialis достигают латеральных ядер таламуса, где начинается третье звено. Аксоны клеток последнего направляются
Рис. 351. Пути трехчленной (а) и двучленной (б) проприо- цептивных рефлекторных дуг.
 1 — мышечные волокна; 2 — двигательные окончания; 3 — сухожилие; 4 — чувствительное окончание в сухожилии;
1 — мышечные волокна; 2 — двигательные окончания; 3 — сухожилие; 4 — чувствительное окончание в сухожилии;
5 — периферическое волокно чувствительного нейрона; 6 — двигательное волокно; 7 — передний корешок; 8 — спинномозговой узел; 9 — задний корешок; 10 — восходящий чувствительный путь заднего канатика; 11 — нисходящий двигательный путь (боковой пирамидный путь); 12 — двигательная клетка переднего рога; 13 — вставочный нейрон.
через capsula interna (см. рис. 297) в кору предцентральной извилины, где помещается корковый конец двигательного анализатора (поля
4, 6; см. рис. 299). По проприоцептивным путям (пройдя спинномозговые нервы) в кору головного мозга поступают нервные импульсы: по fasciculus gracilis — от г мускулатуры нижних конечностей и нижней половины туловища и по fasciculus cuneatus — от верхней половины туловища и верхней конечности. Проприоцептивные волокна от мышц головы проходят по черепным нервам: тройничному (V) — от мышц глаза и от жевательной мускулатуры, VII — от мимической мускулатуры, IX, X, XI и XII — от языка, от мускулатуры глотки и других мышц бывшего жаберного аппарата.
При выпадении глубокой (проприоцептивной) чувствительности больной утрачивает представление о положении частей своего тела в пространстве и о перемене положения; движения теряют свою четкость, согласованность, наступает расстройство координации движений — атаксия. В отличие от мозжечковой (двигательной) атаксии она называется сенсорной (чувствительной).
Не все пути проприоцептивной чувствительности доходят до коры. Подсознательные проприоцептивные импульсы направляются в мозжечок, который является важнейшим центром проприоцептивной чувствительности.
Проприоцептивные пути к мозжечку (рис. 352). Чувствительные подсознательные импульсы от аппарата движения (костей, суставов, мышц и сухожилий) достигают мозжечка посредством спинальных, проприоцептивных путей, из которых главнейшие — tractus spinocerebellaris posterior et anterior (см. рис. 270).
1. Tractus spinocerebellaris posterior. Клеточное тело первого нейрона лежит в спинномозговом узле, аксон делится на две ветви, из которых периферическая идет в составе мышечного нерва от рецептора, заложенного в той или иной части аппарата движения, а центральная в составе заднего корешка проникает в задние рога спинного мозга и при помощи своих концевых ветвей и коллатералей разветвляется вокруг nucleus thoracicus. В niicleus thoracicus лежат клетки
Рис. 352. Восходящие и нисходящие пути мозжечка.
 / — tr. spinocerebellaris posterior; 2, 3 — tr. spinocerebellaris anterior; 2 — часть, перекрещивающаяся в среднем мозге; 3 — часть, перекрещивающаяся в спинном мозге; 4 — tr. rub- rospinalis; 5 — tr. vestibulospinalis; 6 — tr. olivo- spinalis; 7 — tr. thalamocorticalis; S — thalamus;
/ — tr. spinocerebellaris posterior; 2, 3 — tr. spinocerebellaris anterior; 2 — часть, перекрещивающаяся в среднем мозге; 3 — часть, перекрещивающаяся в спинном мозге; 4 — tr. rub- rospinalis; 5 — tr. vestibulospinalis; 6 — tr. olivo- spinalis; 7 — tr. thalamocorticalis; S — thalamus;
9 — nucl. ruber; 10 — червячок мозжечка; 11 — nucl. dentatus cerebelli; 12 — олива и ядра pars vestibularis VIII пары черепных нервов;
13 — связи мозжечка и красного ядра; 14 — tr. corticopontOcerebellaris.
второго нейрона, аксоны которых и образуют tractus spinocerebellaris posterior. Nucleus thoracicus, как показывает название, лучше выражено в грудном отделе на уровне от последнего шейного сегмента до II поясничного. Дойдя в составе бокового канатика спинного мозга до продолговатого по своей стороне, этот тракт в составе нижних мозжечковых ножек достигает коры червя. На своем пути в спинном и продолговатом мозге он не перекрещивается, отчего его называют прямым мозжечковым трактом. Однако, войдя в мозжечок, он большей своей частью перекрещивается в черве.
2. Tractus spinocerebellaris anterior. Первый нейрон тот же, что и у заднего тракта. В substantia intermedia centralis серого вещества спинного мозга помещаются клетки вторых нейронов, аксоны которых, образуя tractus spinocerebellaris anterior, входящий в передние отделы бокового канатика своей и противоположной стороны через commissura alba, совершают в ней перекрест. Тракт поднимается через продолговатый мозг и мост до верхнего мозгового паруса, где снова происходит перекрест. После этого волокна входят в мозжечок через его верхние ножки, где заканчиваются в коре червя. В результате весь этот путь оказывается перекрещенным дважды; вследствие этого проприоцептивная чувствительность передается на ту же сторону, с которой поступила.
Таким образом, оба мозжечковых пути соединяют одноименные половины спинного мозга и мозжечка.
Мозжечок получает также проприоцептивные импульсы от nucleus gracilis и nucleus cuneatus, расположенных в продолговатом мозге. Отростки клеток, заложенных в этих ядрах, идут в мозжечок через его нижние ножки. Все пути глубокой (подсознательной) чувствительности заканчиваются в черве, т. е. в древней части мозжечка, paleocerebellum.
Интероцептивный анализатор
Интероцептивный анализатор в отличие от других не имеет компактной и морфологически строго очерченной проводниковой части, хотя он и сохраняет специфичность на всем своем протяжении.
Рецепторы его, называемые интероцепторами, рассеяны во всех органах растительной жизни; внутренностях, сосудах, в непроизвольной мускулатуре и железах кожи и др.
Кондуктор состоит из афферентных волокон вегетативной нервной системы, идущих в составе симпатических, парасимпатических и анимальных нервов и далее в спинном и головном мозге до коры. Часть кондуктора интероцептивного анализатора составляют афферентные волокна, идущие в составе черепных нервов (V, VII, IX, X) и несущие импульсы от органов растительной жизни, расположенных в области распространения иннервации каждого из этих нервов. Образуемый ими афферентный путь разбивается на 3 звена: клетки первого звена лежат в узлах этих нервов (ganglion trigeminale, ganglion genicuii, ganglion inferius); клетки второго нейрона находятся в ядрах этих нервов (nucleus spinalis n. trigemini, nucleus solitarius nn. VII, IX, X). Исходящие из этих ядер волокна переходят на другую сторону, направляясь к таламусу. Наконец, клетки третьего звена заложены в таламусе.
Значительную часть кондуктора интероцептивного анализатора образует блуждающий нерв, являющийся главным компонентом парасимпатической иннервации. Идущий по нему афферентный путь также разбивается на 3 звена: клетки первых нейронов лежат в ganglion inferius n. vagi, клетки вторых нейронов — в nucleus solitarius.
Исходящие из этого ядра волокна блуждающего нерва вместе с отростками вторых нейронов языкоглоточного нерва переходят на противоположную сторону, перекрещиваясь с волокнами противоположной стороны, и поднимаются по стволовой части мозга. На уровне верхних холмиков крыши среднего мозга они присоединяются ко вторым нейронам кожного анализатора (lemniscus medialis) и достигают таламуса, где лежат клетки третьих нейронов. Отростки последних идут через заднюю треть задней ножки внутренней капсулы к нижнему отделу постцентральной извилины.
В этом месте располагается одна из частей коркового конца интероцептивного анализатора, связанного с парасимпатическими волокнами черепных нервов и областью их иннервации.
Афферентные пути от органов растительной жизни идут также в составе задних корешков спинномозговых нервов. Клетки первых нейронов в этом случае лежат в спинномозговых узлах. Мощный коллектор афферентного пути от органов растительной жизни проходит через внутренностные нервы (nn. splanchnici major et minor). Различные группы нервных волокон этих нервов восходят в спинном мозге в составе его задних и боковых канатиков. Афферентные волокна задних канатиков передают интероцептивные импульсы, достигающие через таламусы коры большого мозга.
Дата добавления: 2016-06-24; просмотров: 582;