ОСНОВЫ БИОЛОГИИ РЫБ 26 страница

|
| -6 |
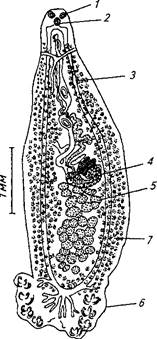
| Рис. 56. Возбудитель дактилогироза Dactylogyrus vastator (из Быховского, 1933): |
| -7 '8 |
Патогенез и симптомы болезни. Под влиянием механического и токсического воздействия гельминтов разрушается или разрастает-
I
1— глаза; 2— глотка; 3 — копулятивный орган; 4— кишечник; 5—яичник; 6— семенник; 7—краевые крючья; 8— срединные крючья; Р —яйцо; 10—личинка; и-— поражение жабр
ся эпителий жаберных лепестков, усиливается отделение слизи, образуются участки некроза, что приводит к нарушению кровообращения и газообмена в жабрах.
Течение инвазии тесно связано с интенсивностью заражения рыб. Больная рыба малоподвижна, скапливается в местах притока свежей воды, богатой кислородом, или держится у ее поверхности, заглатывая воздух. Рыба истощена, глаза западают, жабры обильно покрыты слизью, местами некротизированы и нередко дополнительно заражены грибом сапролегния. В результате гипоксии и асфиксии рыбы погибают.
Диагностика. Учитывают эпизоотологические данные, симптомы болезни и результаты микроскопии соскобов слизи с жаберного аппарата.
Лечение, профилактика и меры борьбы. При дактилогирозах лечение и меры борьбы в основном те же, что и при гиродактилезах. Производителей карпа перед нерестом с профилактической целью обрабатывают в солевых ваннах и после нереста сразу удаляют из нерестовых прудов.
Санитарная оценка. Санитарная оценка при дактилогирозах рыб идентична таковой при гиродактилозах.
Дискокотилез лососевых и хариусовых
Дискокотилез рыб вызывается плоскими гельминтами из класса Monogenea.
Локализация. Взрослые гельминты паразитируют на жабрах рыб.
Распространение и экономический ущерб. Дискокотилез распространен в естественных водоемах северных районов и Сибири, реже — в прудовых хозяйствах Российской Федерации.
Среди зараженной форели нередко отмечаются малокровие и гибель.
Возбудители. У лососевых и хариусовых рыб паразитируют два вида моногенетических сосальщиков, относящихся к семейству Discocotylidae, роду Discocotyle: D. sagittata и D. salmonis. Тело D. sagittata ланцетовидной формы, сплющенное, длиной 6—9 мм, шириной 0,3—0,5 мм. На переднем конце тела имеются две небольшие присоски, на заднем — прикрепительный диск с восемью хитиновыми ущемляющими застежковидными аппаратами, расположенными в два ряда (по 4 аппарата в каждом ряду). Кишечник делится на два ствола, слепо заканчивающихся в заднем конце тела. Половое отверстие находится позади ротового. У них много семенников, расположенных по оси тела в задней половине тела; впереди них находится яичник (рис. 57).
Впереди яичника расположена матка, содержащая одно или несколько яиц. По бокам тела находятся сильно развитые желточни- ки. Тело паразита серовато-бурого цвета.
Биология развития. Паразиты развиваются прямым путем без участия промежуточных хозяев. Гельминты в местах локализации (жабры) выделяют наружу яйца, в которых либо на жабрах, либо в
Рис. 57. Возбудитель дискокотилеза Discocotyle sagittata:
1 — присоски; 2 — глотка; J—кишечник; яичник; 5— семенники; 6— прикрепительный диск; 7—жел- точники
воде развиваются эмбриональные личинки. При температуре воды 18—20 °С в яйцах личинка формируется за 8— 10 сут, при 10—12 °С — за 15—17, а при 5—7 °С — за 20—25 сут. При более низкой температуре развитие личинки в яйце не происходит. Вылупившиеся из яиц личинки прикрепляются в жаберном аппарате дефинитивных хозяев — нельмы, форели, сига, пеляди, муксуна, хариуса и других рыб и через несколько суток превращаются во взрослого гельминта.
Эпизоотологические данные. В России (в водоемах северных районов, Сибири, Карелии, Ленинградской обл.), на Украине (в Закарпатской обл.) дис- кокотилы нередко поражают жаберный аппарат многих ценных рыб семейств Сиговые и Хариусовые. Инвазия встречается в рыбоводных хозяйствах стран Западной Европы и Северной Америки. Заражаются возбудителями как молодь, так и взрослые рыбы. Однако заболевание тяжело, с клиническими признаками протекает у молоди форели и др. Пик инвазии приходится на лето, так как высокая температура воды способствует развитию гельминтов. Взрослые рыбы хотя и заражаются гельминтами, но симптомы болезни проявляются крайне редко. Тем не менее рыбы старших возрастов являются носителями инвазии.
Патогенез и симптомы болезни. Дискокотилы глубоко проникают в жаберные лепестки рыб своим фиксаторным органом. На одной форели интенсивность инвазии достигает 100 экз. и более. Считают, что гельминт способен поглощать значительное количество крови из жабр, в результате чего жабры сильно бледнеют, появляются язвы, кровоизлияния и развивается выраженная анемия.
У больных рыб жабры бледной окраски, покрыты толстым слоем слизи. В местах фиксации гельминтов образуются ранки, которые инфицируются сапрофитной микрофлорой, приводящей к образованию кровоточащих ран. Больные рыбы худеют, отстают в росте. Они часто поднимаются на поверхность воды и заглатывают воздух. Поверхность их тела приобретает пятнисто-матовую окраску.
|
Патологоанатомические изменения. При вскрытии во внутренних органах погибших рыб изменений в основном не находят.
Иногда обнаруживают жировое перерождение мышцы сердца и разрушение плавников. Трупы рыб малокровные, с потускневшей кожей.
Диагностика. Диагноз ставят на основании комплексных эпизо- отологических, клинических и лабораторных исследований с целью обнаружения дискокотилов в соскобах из жабр рыб.
Лечение не разработано.
Профилактика и меры борьбы. В неблагополучных рыбоводных хозяйствах проводят ветеринарно-санитарные и рыбоводно-мели- оративные мероприятия, направленные на предотвращение проникновения возбудителя в водоем и на создание в прудах и бассейнах оптимальных зоогигиенических условий.
Зараженную рыбу обрабатывают 5 мин в противопаразитарных ваннах с 2,5—5,0%-ным раствором поваренной соли. Пруды дезинфицируют хлорной или негашеной известью. При транспортировании рыб из неблагополучного хозяйства ее обязательно проводят через солевые растворы.
Санитарная оценка рыбы. Рыбу, пораженную этой инвазией, если она соответствует товарной кондиции, реализуют без ограничений.
Тетраонхоз сиговых
Тетраонхоз — заболевание рыб, вызываемое плоскими гельминтами из класса Monogenea.
Локализация. Это эктопаразиты рыб, паразитирующие на жаберном аппарате. В естественных водоемах Сибири, Башкортостана и Южного Урала инвазия представлена достаточно широко, где заражаются в основном взрослые сиговые рыбы. При сильной инвазии рыб, особенно осложненной грибами, много продукции выбраковывают из-за потери товарного вида.
Возбудитель. У сиговых рыб паразитирует один вид Tetraonchus alascensis из семейства Tetraonchidae. У щук на жабрах нередко обнаруживают другого возбудителя того же рода — Tetraonchus rnonenteron.
Тело гельминта плоское, достигающее длины 2,0—2,5 мм, ширины 0,6 мм. На заднем конце его имеется прикрепительный аппарат с 16 краевыми и двумя парами серединных крючьев и одной соединительной пластиной. Кишечник в виде одиночного ствола, без боковых выростов, заканчивается слепо. По оси тела в его середине находится округлый семенник. Яичник расположен впереди семенника (рис. 58).
Биология развития. Развитие тетраонхусов происходит прямым путем без участия промежуточных хозяев. В местах локализации гельминты откладывают яйца, из которых в зависимости от температуры воды в разные сроки вылупляются личинки. Последние прикрепляются к жаберному аппарату того же хозяина или других рыб и вскоре достигают половозрелой стадии.
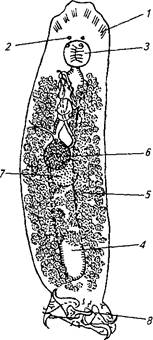
Рис. 58. Возбудитель тетраонхоза Tetraonchus alascensis:
1 — железистые органы; 2—глаза; 5 —глотка; 4—кишечник; 5—желточниют; 6—яичник; 7—семенник; 8— прикрепительный диск
Эпизоотологические данные. В местах распространения гельминта в основном заражаются пыжьян, чир, пелядь, нельма и другие сиговые. Заболевание регистрируется у рыб старших возрастных групп. Энзоотии тетраонхо- зов в основном регистрируют с конца июля по октябрь. Экстенсивность заражения рыб достигает 75—90 % при высокой интенсивности инвазии.
Патогенез и симптомы болезни. Тет- раонхусы, паразитируя в жаберном аппарате, прикрепляются к нему своим мощным прикрепительным органом и, питаясь кровью, разрушают ткани жаберных лепестков. При неблагоприятных условиях эти гельминты могут передвигаться по поверхности рыбы и вновь прикрепляться в разных местах жабр. В водоемах с антисанитарными условиями среды заболевание нередко осложняется поражением жаберных лепестков грибами рода Saprolegnia. На жабрах регистрируют очаги кровоизлияний и некрозы. Все это нарушает нормальный обмен кислорода, и больная рыба проявляет признаки асфиксии. Болезнь нередко принимает тяжелую форму и сопровождается гибелью больных рыб.
Патологоанатомические изменения во внутренних органах не изучены.
Диагностика. Диагноз ставят на основании эпизоотологических, клинических и лабораторных исследований. Обращают внимание на возрастные особенности проявления болезни и время года. Для окончательного уточнения диагноза исследуют жаберные лепестки компрессорным методом или слизь из пораженных участков под малым увеличением микроскопа (х 40 или х 56).
Лечение не разработано.
Профилактика и меры борьбы. В неблагополучных хозяйствах проводят ветеринарно-санитарные, рыбоводно-мелиоративные и биотехнологические мероприятия, направленные на формирование и регулирование стада рыб, не восприимчивых к инвазии. Кроме того, рыбу завозят из благополучных рыбоводных хозяйств.
|
Санитарная оценка рыбы. Возбудитель тетраонхоза не представляет опасности для других животных и человека. Поэтому в зависи
мости от товарного качества рыбу допускают в пищу на общих основаниях либо направляют на рыбозавод для приготовления консервов или кормовой муки ддя животных.
Нитцшиоз осетровых
Нитцшиоз рыб обусловливается паразитированием плоских гельминтов из класса Monogenea.
Локализация. Моногеней паразитируют на лепестках жаберного аппарата, в ротовой полости, прикрепляясь к губам, нёбу.
Распространение и экономический ущерб. Нитцшиоз зарегистрирован у рыб Каспийского и Азовского морей, а также водоемов Западной Европы. Экономический ущерб складывается из потерь в результате уменьшения массы больных рыб, отставания их роста и развития, а также гибели при интенсивной инвазии.
Возбудитель. Nitzschia sturionis относится к семейству Capsa- lidae. Тело гельминта продолговато-овальной формы, длиной 10— 25 мм, шириной 3,2 мм. На переднем конце тела имеются две железистые, продольно сжатые присоски. Два толстых кишечных ствола с боковыми отростками в задней части тела заканчиваются слепо. Семенников много, они овальной формы, расположены по средней линии в задней части гельминта. Яичник расположен впереди семенников. Половое отверстие открывается в средней части тела, позади глотки (рис. 59).
Биология развития. Развитие нитц- шии происходит прямым путем. Как и многие моногенетические гельминты, половозрелые нитцшии откладывают яйца на жабрах и в ротовой полости рыб, которые с помощью липкого секрета прикрепляются в местах их пара- зитирования. Из яиц вскоре вылупляются личинки, способные прикрепляться к жабрам и слизистой оболочке ротовой полости хозяина, или попадают в воду и поселяются на других особях, где они вскоре достигают стадии имаго. В воде личинки способны заражать своего хозяина в течение 5—6 ч.
Взрослые гельминты после гибели зараженной рыбы живут не более 24 ч.
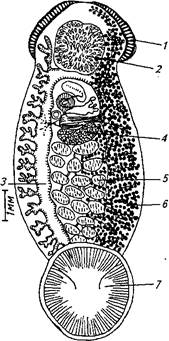
Рис. 59. Возбудитель нитцшиоза Nitzschia sturionis:
|
1 — железистый орган; 2— присоска; 3 — кишечник; яичник; 5—семенники; 6 — желточники; 7— прикрепительный диск
Эпизоотологические данные- К инвазии восприимчивы осетровые рыбы: осетр, севрюга, шип и др. Заражаются как взрослые особи, так и молодь рыб. Источником распространения инвазии служат больные рыбы. Пик заболевания чаще приходится на весенне-летний период. В осенне-зимний сезон у рыб отмечают па- разитоносительство. Паразиты развиваются только в морской воде. В период нерестовых миграций осетровых рыб в пресные воды нитцший у них не обнаруживают. Реже они встречаются в сильно опресненных участках морских акваторий. Согласно наблюдениям интенсивность инвазии у шипа достигала 150— 600 экз.
Патогенез и симптомы болезни. Нитцшии в местах паразитирова- ния питаются кровью, поглощая ежесуточно около 0,5 см3. При значительном количестве гельминтов возникает воспаление в жаберных лепестках и слизистых оболочках ротовой полости. В результате нарушения кровоснабжения вторичные лепестки атрофируются и вместе с окружающими тканями жабр подвергаются некрозу. Нарушаются кровообращение и газообмен. У рыб развивается выраженная анемия. Они худеют, замедляются рост и развитие молоди, что нередко приводит к их гибели.
Патологоанатомические изменения. Они изучены недостаточно. У молоди рыб жаберные лепестки представляют собой сплошную белую массу, состоящую из соединительной ткани. Погибшие рыбы сильно истощены.
Диагностика. На основании исследований эпизоотологии и симптомов болезни ставится предварительный диагноз, который подтверждается микроскопированием слизи из жаберного аппарата и ротовой полости на наличие гельминтов.
Лечение не разработано.
Профилактика и меры борьбы. Основное внимание уделяется профилактическим мерам, направленным на предотвращение заноса возбудителя из неблагополучных водоемов в благополучные, где имеются восприимчивые к нему виды рыб. С этой целью рыбу, предназначенную для перевозки в другие водоемы, тщательно исследуют
В 1939 г. каспийская севрюга была завезена в Аральское море с нарушением ветеринарно-санитарных правил, в результате чего у аральского шипа нитцшиоз достиг за короткий срок высокой экстенсивности и интенсивности заражения и вызвал массовую гибель рыб.
Санитарная оценка рыбы. Для человека и животных данный гельминт не представляет опасности. Поэтому, если рыба, пораженная нитцшиями, сохранила товарную ковдицию, ее допускают в пищу людям без ограничений; рыбу, не соответствующую товарной кондиции, при высокой интенсивности заражения, с признаками истощения по усмотрению специалистов направляют на корм плотоядным животным и птице.
Диплозоонозы пресноводных рыб
Диплозоонозы — заболевания многих видов пресноводных и реже морских рыб, вызываемые плоскими гельминтами из класса Monogenea.
Локализация. Гельминты паразитируют, как и многие другие моногеней, в жаберном аппарате рыб.
Распространение и экономический ущерб. Возбудители диплозоо- нозов широко распространены в реках и озерах, относящихся к бассейнам Балтийского, Азовского, Каспийского и Черного морей, а также в реках Амуре, Енисее, Лене и Волге.
Экономический ущерб складывается из потерь массы больной рыбы.
Возбудители. В последнее время установлено более 15 видов диплозоон, паразитирующих у различных рыб. Так, на жабрах плотвы паразитирует Diplozoon homoion, у язя — D. megan, у пескаря — D. gracilis, у белоглазки — D. bergi, у усача — D. tatzhikista- nicum, у маринки — D. schizothorazi и др. Поскольку все они по многим параметрам идентичны, то мы даем полное описание только одного гельминта — Diplozoon paradoxum. Все указанные выше виды относятся к семейству Discocotylidae, роду Diplozoon.
Это достаточно крупные плоские гельминты длиной 4—12 мм и шириной 0,35—0,53 мм. Свежие паразиты имеют грязно-серый цвет с коричневым оттенком. Передний конец тела снабжен небольшой присоской, на дне которой расположено ротовое отверстие. В заднем конце тела гельминтов находится прикрепительный (фиксаторный) аппарат, состоящий из хитиновых образований, напоминающих пряжки. Количество прикрепительных аппаратов у разных видов диплозоонов различное, что имеет большое диагностическое значение.
Диплозооны обладают чрезвычайно характерным для них свойством, отразившимся на их биологии и морфологии. Гельминты в преимагинальной стадии обитают в жаберном аппарате в отдельности, но, достигнув стадии имаго, две особи сходятся вместе и срастаются в области брюшной присоски. При этом женская половая система одного гельминта срастается с мужской другого. Два сросшихся гельминта приобретают Х-образную форму (рис. 60).
Биология развития. Развитие гельминтов происходит прямым путем. Диплозооны паразитируют у леща, густеры, чехони, плотвы, усача, язя, красноперки, шемаи, белоглазки, налима, бычка и др. Паразиты в местах локализации откладывают яйца, у которых на одном полюсе имеется длинная спиралеобразная филамента. Из яиц вылупляются личинки (дипорпа), у которых постепенно развиваются три пары присасывательных клапанов. Личинки прикрепляются к лепесткам жабр и вырастают до определенной стадии своего развития. Затем к моменту достижения половой зрелости они попарно сходятся, срастаются в области брюшной присоски и далее образуется спайник.
Рис. 60. Возбудитель диплозооноза Diplozoon paradoxum: 1 — присоска; 2 — глотка; 3 — кишечник; 4 — прикрепительные диски с крючьями
Эпизоотологические данные. Диплозоонозы — заболевания пресноводных рыб, как речных, так и прудовых, разводимых в рыбоводных хозяйствах. Источниками распространения инвазии являются больные рыбы. Диплозооны паразитируют у рыб разных возрастных групп, однако наибольшая зараженность отмечается у рыб в 2-летнем возрасте.
Экстенсивность инвазии достигает 75—85 % при интенсивности до 17—38 экз. и более. В заиленных и слабопроточных водоемах экстенсивность и интенсивность заражения рыб, как правило, бывают выше.
Патогенез и симптомы болезни. Своими мощными прикрепительными органами гельминты сильно травмируют жаберный аппарат. В местах разрушения лепестков разрастается соединительная ткань, затрудняющая газообмен. На жабрах обнаруживают припухлости, сплошь покрытые густой слизью. При обострении патологического процесса на жабрах развивается сапролегния, которая усугубляет течение болезни. Рыба постепенно слабеет, худеет; для нее характерна ограниченная подвижность.
Патологоанатомические изменения. Они описаны недостаточно, но тем не менее заметны поражения жаберного аппарата. Группы погибших рыб истощены, жабры и рот раскрыты, глаза мутные.
Диагностика. Диагноз ставится на основании исследования под малым увеличением микроскопа слизи с поверхности жаберного аппарата и жаберных лепестков. При диагностике диплозоонозов учитывают и эпизоотологические данные, а также симптомы болезни.
Лечение не разработано.
Профилактика и меры борьбы. Для снижения зараженности рыб диплозоонами в прудовых хозяйствах выростные и нагульные пруды содержат в хорошем состоянии, соответствующем ветеринарно- санитарным требованиям. По возможности пруды дезинвазируют и просушивают после отлова рыбы. Весной для удаления ила и уничтожения яиц и личинок гельминтов ложе пруда перепахивают. В естественных водоемах проводят мелиоративные работы.
При обнаружении гельминтов в прудовых хозяйствах карпов купают в солевых ваннах, как и при других моногенеидозах.
Санитарная оценка рыбы. Так как диплозооны не представляют опасности для человека, то зараженную рыбу, соответствующую требованиям товарной кондиции, реализуют в пищу без ограничений.
Трематодозы
Систематика и краткая характеристика трематод. Трематодозы — инвазионные заболевания, возбудителями которых являются различные виды трематод или их личинки, относящиеся к классу трематод — Trematoda, или дигенетических сосальщиков, типу Plathelminthes.
Анатомия и биология трематод. Это плоские черви листовидной формы. Тело обычно удлиненно-овальное, размером от 1 мм до 15 см. Оно покрыто тегументом (гладкое, с шипами или с щетинками) , состоящим из наружного и внутреннего слоев (рис. 61). У большинства трематод две присоски: ротовая (на переднем конце тела) и брюшная (чаще на середине тела). У некоторых трематод, например у представителей Sanguinicolidae, паразитирующих в крови рыб, присоски рудиментированы или отсутствуют. От ротового отверстия отходит глотка, затем пищевод, переходящий в кишечник. Последний в виде двух стволов, достигнув заднего конца тела, слепо заканчивается. В очень редких случаях кишечные стволы открываются наружу. Сосальщики питаются тканевой жидкостью, слизью, некоторые — кровью. Продукты обмена выделяются через экскреторную систему протонефридиального типа. По всему телу размещены канальцы, объединяющиеся в два выделительных канала, открывающихся на заднем конце тела.
Нервная система состоит из парного надглоточного ганглия и отходящих нервных стволов.
Дигенетические сосальщики рыб — гермафродиты.
Мужская половая система представлена в основном двумя семенниками (реже больше), от которых отходят семяпроводы, впадающие в общий семявыносящий проток. Последний образует не-
А ЦМ Ш БС В Б
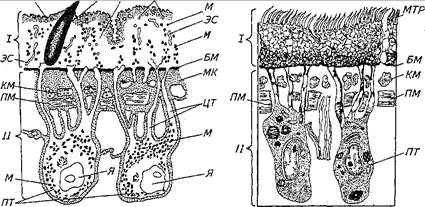 Рис. 61. Схема строения покровов (тегумента) трематод (А), дестод (Б) по результатам электронной микроскопии (из Р. С. шульца, Е. В. Гвоздева, 1970):
/—наружная часть: ЦМ — цитоплазматйческая мембрана; Ш —шипик; БС — безъядерный синцитий; В — вакуоли; M — митохондрии; ЭС — эндоплазматическая сеть; БМ — базальная мембрана; II— внутренняя часть: КМ — кольцевые мышцы; ПМ — продольные мышцы; MK — межклеточное вещество; ЦТ-—щгго плазматические тяжи; Я— ядра; ПТ — погруженная часть тегумента; МТР — микротрихии
Рис. 61. Схема строения покровов (тегумента) трематод (А), дестод (Б) по результатам электронной микроскопии (из Р. С. шульца, Е. В. Гвоздева, 1970):
/—наружная часть: ЦМ — цитоплазматйческая мембрана; Ш —шипик; БС — безъядерный синцитий; В — вакуоли; M — митохондрии; ЭС — эндоплазматическая сеть; БМ — базальная мембрана; II— внутренняя часть: КМ — кольцевые мышцы; ПМ — продольные мышцы; MK — межклеточное вещество; ЦТ-—щгго плазматические тяжи; Я— ядра; ПТ — погруженная часть тегумента; МТР — микротрихии
|
большое расширение — семенной пузырек. Конечный отдел се~ мяпротока — семяизвергательный канал. Имеется мышечный ко- пулятивный орган, помещающийся в циррусном мешке.
Женская половая система представлена яичниками различной формы, от которых отходит яйцепровод, открывающийся в небольшое мешковидное образование — оотип. В нем происходят формирование яиц и оплодотворение. По протокам в оотип поступают желточные клетки из желточников, размещенных по бокам тела. Протоки сливаются в общий канал. В оотип открываются также семяприемник и тельце Мелиса, выделяющее особый секрет. Матка имеет вид извитого расширенного канала, открывающегося наружу возле мужского полового отверстия в общий половой синус или клоаку. Половой синус обычно расположен между ротовой и брюшной присосками. Созревшие яйца из матки выводятся наружу через наружное половое отверстие.
Биология развития. Развитие дигенетических сосальщиков происходит со сменой нескольких поколений личинок и со сменой двух или трех хозяев. Эмбриональное развитие оплодотворенных яиц происходит в матке. Когда яйцо попадает в воду, в нем формируется личинка, покрытая ресничками (мирацидий), которая выходит из яйца в воду. Мирацидий, плавая в воде, проникает в первого промежуточного хозяина, а им, как правило, является пресноводный моллюск. В печени моллюска совершается бесполое размножение личинки. Мирацидий превращается в мешковидный организм — спороцисту. Внутри спороцисты из зародышевых клеток развиваются редии, которые дают начало церкариям, несколько напоминающим взрослых трематод. Характерным для церкариев является наличие хвоста различной длины и формы. Сформировавшиеся церкарии выходят из тела моллюска и некоторое время плавают в воде. Вторым (дополнительным) хозяином является рыба. Церкарии внедряются в тело рыбы и поселяются в различных органах и тканях: под кожей, в мышцах, хрусталике или стекловидном теле глаз и др. В теле дополнительного хозяина формируется инвазионная личинка — метацеркарий. Иногда личинки инцистируют- ся, вокруг них образуется соединительнотканная оболочка. Мета- церкарии в теле рыб могут сохраняться 2—3 года. В отличие от взрослого гельминта у метацеркарйя отсутствует лишь развитая половая система. Для достижения половой зрелости метацеркариям необходимо попасть в организм окончательных (дефинитивных) хозяев. В зависимости от вида трематод ими являются различные виды рыбоядных птиц (чайки, крачки, крохали, цапли, пеликаны, бакланы и др.), плотоядные животные и человек. В организме окончательного хозяина метацеркарии за несколько дней превращаются во взрослую стадию паразита (имаго) и начинают выделять яйца. На этом биология развития замыкается. Скорость развития личинок во внешней среде и партеногенетических поколений в моллюсках зависит от факторов водной среды. Главную роль в этом играет температурный режим. В весенне-летний период, когда температура воды достигает 20—24 °С, личинки развиваются гораздо быстрее, чем в осеннее время, при более низкой температуре. Зимой развитие или прекращается, или продолжается в течение более длительных сроков.
У рыб трематодозы чаще вызываются личиночными стадиями гельминтов — церкариями и метацеркариями. Лишь небольшое количество сосальщиков в половозрелой стадии паразитирует у рыб, например Sanguinicola inermis.
Класс Trematoda включает сотни видов сосальщиков, паразитирующих у рыб. Однако не все из них имеют патогенное значение. Укажем только те виды, которые вызывают заболевания рыб.
Диплостомоз
Диплостомоз вызывается метацеркариями трематоды из семейства Diplostomidae, класса Trematoda.
Локализация. Места локализации метацеркариев — хрусталик, стекловидное тело, оболочки глаза, головной мозг и реже другие органы рыб.
Распространение и экономический ущерб. Заболевание распространено повсеместно. Заражению подвержены рыбы более 100 видов в естественных водоемах и в прудовых хозяйствах. При высокой интенсивности инвазии рыбы старшего возраста отстают в росте и развитии, а также истребляются хищниками. Гибель личинок и мальков достигает 70—85 %. Личинки погибают от 1—2 метацерка- риев, мальки — от 3—5 метацеркариев.
Возбудитель. Возбудителем является метацеркарий сосальщика — Diplostomum spathaceum. Взрослый паразит плоский, с длиной тела 0,4—0,5 см и шириной 0,2—0,3 см. В середине тела есть перетяжка, которая делит его на переднюю листовидно-расширенную часть и заднюю часть, более узкую, короткую, цилиндрическую. Ротовая присоска и два железистых образования расположены в передней части тела; брюшная присоска находится в середине тела. Округлые яичники, матка и семенники расположены в задней части тела. Яйца овальной формы, с крышечкой.
Метацеркарии овальной формы, длиной 0,3—0,4 мм. На переднем конце имеются два ушковидных выроста и ротовая присоска. Брюшная присоска находится в середине вентральной стороны тела, а за ней каудально расположен железистый орган Брандеса (рис. 62).
Биология развития. Дефинитивными хозяевами являются рыбоядные птицы, преимущественно чайковые, реже — рыбоядные утки. Промежуточные хозяева — пресноводные моллюски Lymneae stagnalis, Radix ovata и др., дополнительные — рыбы (карп, лещ, окунь, щука, форель, толстолобик и др.). В кишечнике птицы гельминты откладывают яйца, которые при выходе наружу содержат мирацидии. Мирацвдии, плавая в воде, отыскивают моллюсков и внедряются в их печень, где происходят бесполое размножение, образование спороцисты, редии и вилохвостых церкариев. Церкарии покидают организм моллюска и, попав в воду, внедряются через кожный покров в мышцы рыб и затем по кровеносным сосудам попадают в глаза, далее в хрусталик, где вскоре превращаются в метацеркариев. Считают, что метацеркарии в рыбе могут быть жизнеспособными 3—4 года. В кишечнике птиц трематоды достигают половой зрелости через 4—5 сут (рис. 63).
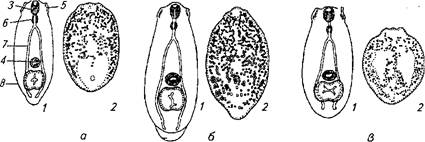 Рис. 62. Возбудители дипдостомоза — метацеркарии (из Шигина, 1968):
а — Diplostomum spathaceum; б— D. megri; в — D. helveticum; 1 — общий вид; 2— расположение известковых телец; 3 — ротовая присоска; брюшная присоска; 5 —ушки; <5— глотка; 7— кишечник; 8— железистый орган Брандеса
Рис. 62. Возбудители дипдостомоза — метацеркарии (из Шигина, 1968):
а — Diplostomum spathaceum; б— D. megri; в — D. helveticum; 1 — общий вид; 2— расположение известковых телец; 3 — ротовая присоска; брюшная присоска; 5 —ушки; <5— глотка; 7— кишечник; 8— железистый орган Брандеса
|
Рис. 63. Биология развития Diplostomum spathaceum (из Бауера и др., 1981):
1 — взрослый паразит; 2— яйцо; 3 — мирацвдий; 4 — спороциста, редии; 5 — церкарий; 6— ме- тацеркариЙ; 7— первый промежуточный хозяин (моллюск); 8— второй промежуточный хозяин (рыба); дефинитивный хозяин (чайка)
Эпизоотологические данные. Ареал диплостомоза чрезвычайно широк. Это заболевание встречается среди большого количества видов рыб (карп, лещ, елец, окунь, судак, налим, щука, форель, омуль, белый амур и др.), но более опасен для лососей, форели, сигов, белого и пестрого толстолобиков.
Источником распространения трематоды являются инвазиро- ванные птицы, резервуаром инвазии — брюхоногие моллюски. Большую роль в распространении возбудителя играют рыбоядные птицы, которые, совершая суточные и сезонные миграции, могут
переносить возбудителя болезни на большие расстояния. Заражение рыб и пик диплостомоза чаще наблюдаются в весенне-летний период. Наиболее часто заболевание регистрируют в выростных прудах среди мальков и сеголетков после пересадки молоди из нерестовых прудов. Экстенсивность инвазии составляет 60—100 % при интенсивности 5—25 метацеркариев и более. Болезнь встречается и в нагульных водоемах, хотя более взрослые популяции рыб заражаются не так интенсивно.
Дата добавления: 2016-06-24; просмотров: 817;