Пресинаптическое торможение
Схема опыта открытия пресинаптического торможения приводится на рис. 40.
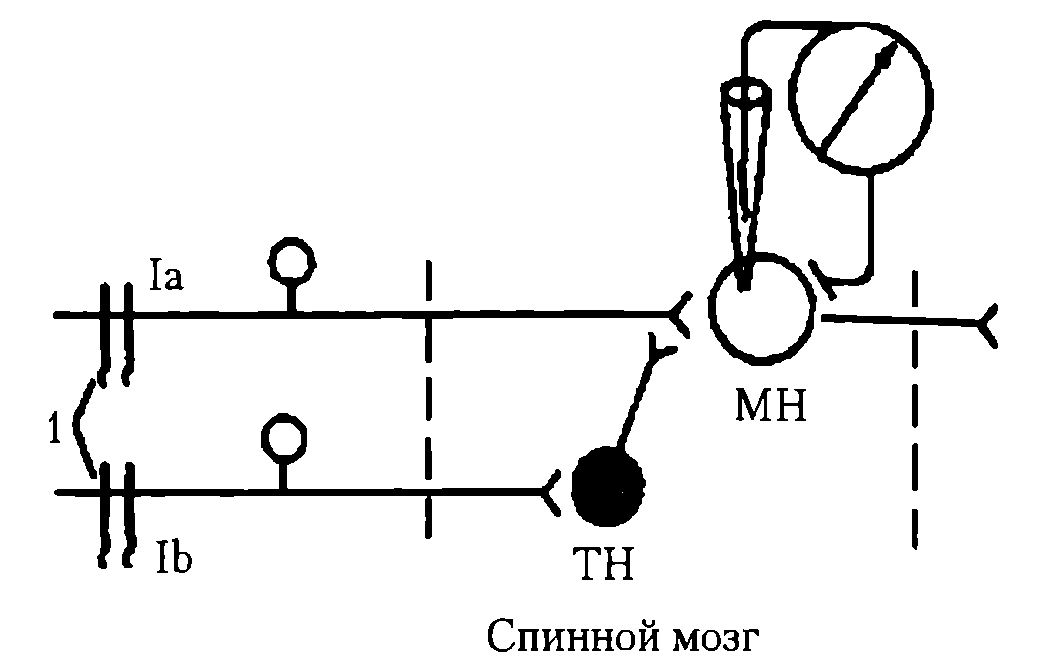
Рис. 40. Схема опыта, доказывающего наличие пресинаптического торможения. При раздражении Ia в МН регистрируется ПД, при опережающем раздражении на 20 мс Ib в МН ПД не возникает: Ia – афферентный путь от мышечных рецепторов мышц-разгибателей (моносинаптическая рефлекторная дуга); Ib – афферентный путь от сухожильных рецепторов мышц-сгибателей; 1 – раздражающие электроды; МН – мотонейрон спинного мозга; ТН – тормозной нейрон
Известно, что раздражение первичных афферентов мышечных веретен сопровождается возбуждением гомонимных α-мотонейронов (α-мотонейронов этой же мышцы). Однако опережающее раздражение афферентов сухожильных рецепторов мышц-антагонистов предотвращает возбуждение активируемых α-мотонейронов. При этом мембранный потенциал и возбудимость исследуемых α-мотонейронов не изменялись либо регистрировался низкоамплитудный ВПСП, недостаточный для возникновения ПД. Поскольку в опыте исследовались мотонейроны в составе моносинаптической рефлекторной дуги, было ясно, что они не возбуждаются вследствие процессов, происходящих в пресинаптическом окончании. Это и определяет название данного вида торможения, так как возбуждение блокируется на подступах к постсинаптическому нейрону.
В основе механизма пресинаптического торможения лежит деполяризация пресинаптических окончаний (тормозная пресинаптическая деполяризация — ТПД). В очаге деполяризации нарушается процесс распространения возбуждения, и, следовательно, поступающие к нервному окончанию импульсы, не имея возможности пройти зону деполяризации в обычном количестве и обычной амплитуды, не обеспечивают выделения медиатора из пресинаптического окончания в синаптическую щель в достаточном количестве — постсинаптический нейрон не возбуждается, его функциональное состояние, естественно, остается неизменным. Деполяризацию пресинаптической терминали вызывают специальные тормозные вставочные клетки, аксоны которых образуют синапсы на пресинаптических окончаниях аксона мишени (рис. 41).
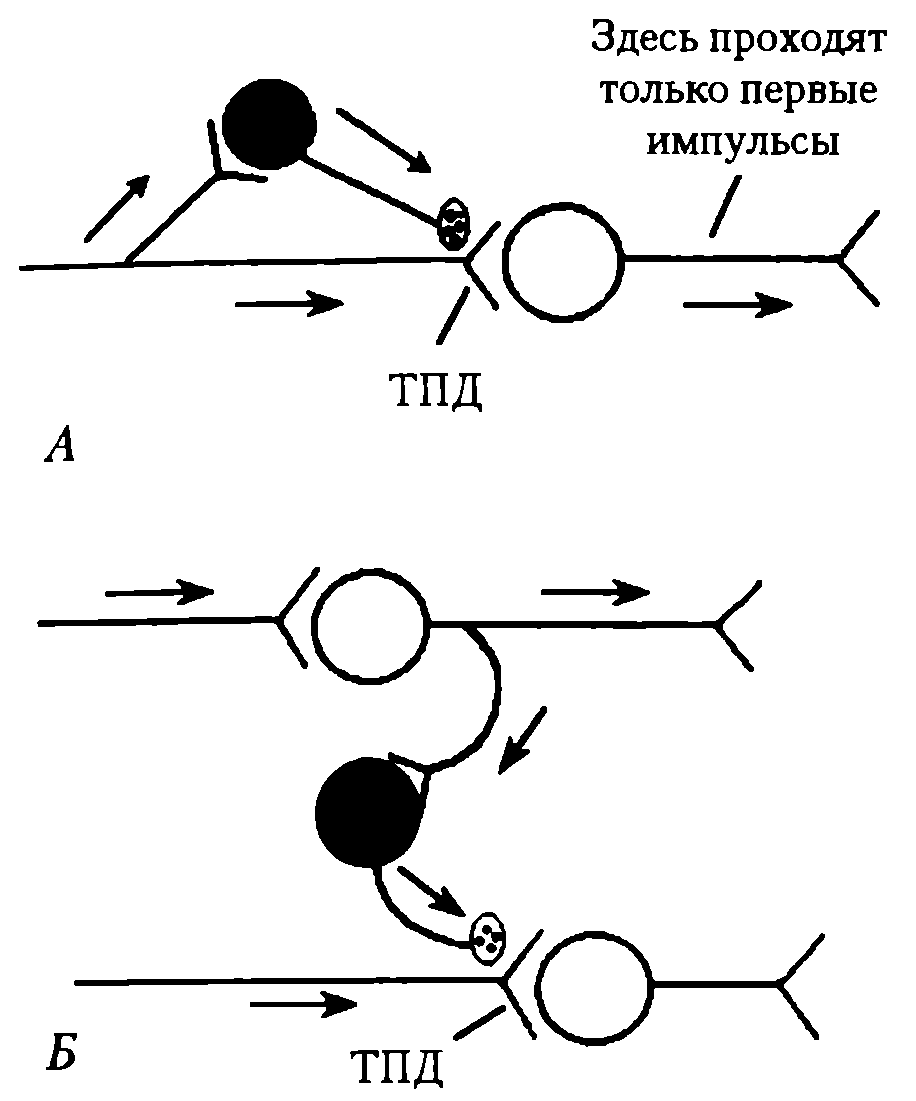
Рис. 41. Разновидности пресинаптического торможения: А – параллельное; Б – латеральное. Нейроны: светлые – возбуждающие, черные - тормозные
Снижение ПД в пресинаптической терминали (частичная деполяризация) всего лишь на 5% уменьшает ВПСП на постсинаптической мембране на 50%.
Торможение (деполяризация) терминали после одного афферентного залпа продолжается 300—400 мс; медиатором служит ГАМК, которая действует на ГАМК1-рецепторы.
ТПД является следствием повышения проницаемости для С1- и выхода его из клетки. Этот факт свидетельствует о том, что в составе мембран пресинаптических терминалей имеется хлорный насос, обеспечивающий первичный транспорт С1- внутрь клетки вопреки электрическому градиенту. Под действием ГАМК тормозных нейронов и последующего повышения проницаемости мембраны для С1- ионы последнего начинают выходить наружу согласно электрическому градиенту, но вопреки концентрационному. Это приводит к деполяризации пресинаптических терминалей и ухудшению их способности проводить импульсы к постсинаптическому нейрону. Роль ГАМК2-рецепторов на пресинаптических окончаниях изучена недостаточно. Хотя известно, что активация ГАМК2- рецепторов на постсинаптической мембране нейрона ведет к выходу ионов К+ из клетки (гиперполяризация) и снижению возбудимости.
Известны параллельное и латеральное пресинаптическое торможение (см. рис. 39). Возвратное пресинаптическое торможение на уровне спинного мозга (по типу возвратного постсинаптического торможения) у млекопитающих обнаружить не удалось, хотя у лягушек оно выявлено. В реальной действительности взаимоотношения возбуждающих и тормозных нейронов значительно сложнее. Тем не менее, все варианты пре- и постсинаптического торможения можно объединить в две группы: 1) когда распространяющееся возбуждение блокирует собственный путь с помощью вставочных тормозных клеток (параллельное и возвратное торможение) и 2) когда распространяющееся возбуждение блокирует другие нервные пути с помощью включения тормозных клеток (латеральное и прямое торможение). Поскольку тормозные клетки сами могут быть заторможены другими тормозными нейронами, это может облегчить распространение возбуждения, т.е. наблюдается феномен растормаживания.
Иногда в качестве разновидности центрального торможения выделяют торможение вслед за возбуждением.
Исходя из имеющихся фактов, особым механизмом торможения считать его нельзя, поскольку оно может быть результатом следовой гиперполяризации нейронов либо возвратного торможения. В спинном мозге это осуществляется с помощью клеток Реншоу. Пессимальное торможение (пессимум Введенского), наблюдаемое в эксперименте на нервно-мышечном препарате, в ЦНС в физиологических условиях также, по-видимому, не встречается.
Дата добавления: 2016-02-16; просмотров: 6456;