Свойства нервных центров
Свойства нервных центров связаны с некоторыми особенностями распространения возбуждения в ЦНС, особыми свойствами химических синапсов и свойствами мембран нервных клеток.
Нервным центрам свойственно явление суммации возбуждений, открытое И. М. Сеченовым (1868) в опыте на лягушке: раздражение конечности слабыми редкими импульсами не вызывает реакции, а более частые раздражения такими же слабыми импульсами сопровождаются ответной реакцией — лягушка совершает прыжок. Различают временную (последовательную) и пространственную суммации (рис. 36).
Временная суммация показана на рис. 36, А: слева — схема для экспериментального тестирования эффектов, вызываемых в нейроне ритмической стимуляцией аксона; справа — запись, которая позволяет видеть, что если ВПСП быстро следуют друг за другом, то они суммируются благодаря своему относительно медленному временному ходу (несколько миллисекунд), достигая в конце концов порогового уровня. Временная суммация обусловлена тем, что ВПСП от предыдущего импульса еще продолжается, когда приходит следующий импульс. Поэтому данный вид суммации называют также последовательной суммацией. Ее роль связана с тем, что многие нейронные процессы имеют ритмический характер и, таким образом, могут суммироваться, давая начало возбуждению в нейронных объединениях нервных центров.
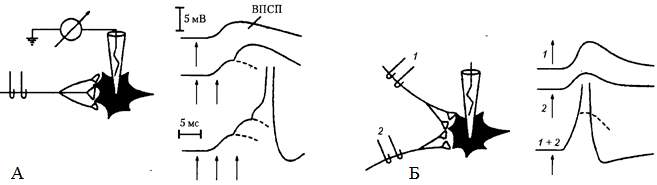
Рис. 36. Суммация возбуждений в нейроне: А – временная: один стимул (↑) и два (↑↑) вызывают подпороговый ВПСП, три последовательных стимула (↑↑↑) обеспечивают возникновение ПД; Б – пространственная суммация: раздельные одиночные раздражения (1 и 2) вызывают подпороговые ВПСП, одновременные два раздражения вызывают ПД
Пространственная суммация проиллюстрирована на рис. 36, Б, где показано, что раздельная стимуляция каждого из двух аксонов вызывает подпороговый ВПСП, тогда как при одновременной стимуляции обоих аксонов возникает ПД. Пространственная суммация связана с такой особенностью распространения возбуждения, как конвергенция, т.е. схождение многих афферентных путей к одному и тому же центру.
Для нервного центра характерно последействие — это продолжение возбуждения нервного центра после прекращения поступления к нему импульсов по афферентным нервным путям. Главной причиной последействия является циркуляция возбуждения по замкнутым нейронным цепям, которая может продолжаться минуты и даже часы. Таким образом, особенность распространения возбуждения (его циркуляция) обеспечивает другое явление в ЦНС — последействие. Последействие играет важнейшую роль в процессах обучения — это начальный этап процесса запоминания информации. Очень краткое последействие (десятки-сотни миллисекунд) объясняется продолжающимся существованием ВПСП, на фоне которого возникает несколько ПД. Таким же кратким последействие может быть при повторном появлении следовой деполяризации, что свойственно нейронам ЦНС. Если следовая деполяризация достигает КП, то возникает ПД. Последействие способствует проявлению следующих свойств нервных центров.
Нервные центры обладают постоянной (фоновой) активностью (тонусом), что объясняется: 1) спонтанной активностью нейронов ЦНС; 2) влияниями БАВ (метаболиты, гормоны, медиаторы и др.), циркулирующих в крови и влияющих на возбудимость нейронов; 3) афферентной импульсацией от различных рефлексогенных зон; 4) циркуляцией возбуждения в ЦНС; 5) суммацией миниатюрных потенциалов, возникающих в результате спонтанного выделения квантов медиатора из аксонов, образующих синапсы на нейронах.
Значение фоновой активности нервных центров заключается в обеспечении некоторого исходного уровня деятельного состояния центра и эффекторов. Этот уровень может увеличиваться уменьшаться в зависимости от колебаний суммарной активности нейронов нервного центра-регулятора и обеспечивает двоякое влияние на деятельность других центров и регулируемого рабочего органа — стимулирующее и угнетающее.
Нервным центрам присуща трансформация ритма возбуждения — изменение числа импульсов, возникающих в нейронах центра на выходе, относительно числа импульсов, поступающих на вход данного центра. Трансформация ритма возбуждения возможна в сторону как увеличения, так и уменьшения. Уменьшение числа импульсов в нервном центре объясняется снижением возбудимости последнего за счет процессов пре- и постсинаптического торможения, а также избыточным потоком афферентных импульсов. При большом потоке афферентных импульсов, когда уже все нейроны центра или нейронного пула возбуждены, дальнейшее увеличение афферентных входов не увеличивает число возбужденных нейронов и нервных импульсов на выходе. Из сказанного видно, что выделение феномена «окклюзия» (по Шеррингтону, «закупорка»; от лат. оссlusum — закрывать) в настоящее время представляется нецелесообразным, несмотря на то, что этот феномен и фигурирует в современной литературе. Увеличению числа импульсов, возникающих в центре в ответ на афферентную импульсацию, способствуют иррадиация процесса возбуждения и последействие.
Важнейшим свойством нервных центров является их пластичность — способность к перестройке функциональных свойств. Пластичность включает ряд феноменов.
Синаптическая потенциация — улучшение проведения в синапсах после кратковременной их активации, которая ведет к увеличению амплитуды постсинаптических потенциалов. Степень выраженности потенциации возрастает с увеличением частоты импульсов; оно максимально, когда импульсы поступают с интервалом в несколько миллисекунд. Длительность синаптической потенциации после одиночных стимулов мала; после раздражающей серии она может составлять от нескольких минут до нескольких часов и даже нескольких дней. Одной из причин возникновения синаптической потенциации является накопление Са2+ в пресинаптических окончаниях, поскольку Са2+, который входит в нервное окончание во время ПД, способствует высвобождению медиатора. Важным фактором является ускорение синтеза рецепторов и медиатора, мобилизация пузырьков при частом использовании синапсов. Напротив, при редком использовании синапсов синтез медиаторов уменьшается — важнейшее свойство ЦНС. Поэтому фоновая активность нейронов способствует возникновению возбуждения в нервных центрах, повышает его функциональные возможности.
В случае длительной синаптической потенциации главную роль играют метаболические процессы, запускаемые в нейронах с помощью метаботропных рецепторов и вторых посредников. В частности, активируются Са-зависимые протеазы, что ведет к эффективности синаптической передачи. Особенно продолжительна синаптическая потенциация в результате активации синтеза нейромодуляторных пептидов (эндорфины, энкефалины, ангиотензин II). Интенсивное функционирование синапсов ведет к миелинизации пресинаптических окончаний и увеличению эффективности передачи, что, по-видимому, связано с меньшей утечкой ионов через неуправляемые каналы (миелин является изолятором).
Значение синаптической потенциации заключается в том, что оно создает предпосылки улучшения процессов переработки информации в нейронах нервных центров, что крайне важно, например, для обучения в ходе выработки двигательных навыков, условных рефлексов, запоминания любой информации. Повторное возникновение явлений потенциации в нервном центре может вызвать переход последнего из обычного состояния в доминантное, характеризующееся рядом новых свойств, которые рассматриваются ниже.
Доминанта (от лат. dominans— господствующий) — это стойкий господствующий очаг возбуждения в ЦНС, подчиняющий себе функции других нервных центров. Явление доминанты открыл А. А. Ухтомский (1923) в опытах с раздражением двигательных зон коры большого мозга, вызывающим сгибание конечности животного. Как выяснилось, если раздражать корковую двигательную зону (например, сгибание нижней конечности) на фоне сильного повышения возбудимости другого нервного центра, сгибания конечности может не произойти. Раздражение двигательной зоны вызывает реакцию того органа, деятельность которого контролируется господствующим, т.е. доминирующим, в данный момент в ЦНС нервным центром, например центром мочеиспускания.
Доминантное состояние нервного центра можно получить многократной посылкой афферентных импульсов к определенному центру, гуморальными на него влияниями. Роль гормонов в образовании доминантного очага возбуждения демонстрирует опыт на лягушке: весной у самца раздражение любого участка кожи вызывает не защитный рефлекс, а усиление обнимательного рефлекса. В условиях натурального поведения доминантное состояние нервных центров может быть вызвано метаболическими сдвигами показателей внутренней среды, например жажда, возникающая при повышении осмоляльности крови.
Главные свойства доминантного очага возбуждения: инерционность, повышенная возбудимость, способность реагировать на возбуждения, иррадиирующие по ЦНС, стойкость возникшего возбуждения, способность оказывать угнетающее влияние на центры-конкуренты и другие нервные центры.
Биологическая роль доминантного очага возбуждения в ЦНС заключается в том, что на его основе формируется конкретная приспособительная деятельность, ориентированная на достижение полезного результата, необходимого для устранения причин, поддерживающих тот или иной нервный центр в доминантном состоянии. Например, на базе доминантного состояния центра голода реализуется пищедобывательное поведение, на базе доминантного состояния центра жажды побуждается поведение, направленное на поиск воды. Доминантные взаимодействия двух возбужденных центров лежат в основе замыкания временной связи при выработке условных рефлексов. Доминантное состояние двигательных центров обеспечивает выполнение автоматизированных реакций (например, работа на конвейере, выполнение комплекса гимнастических упражнений и т.п.).
Синаптическая депрессия (утомляемость нервных центров и восстановление работоспособности после отдыха) также является одним из вариантов пластичности ЦНС. Утомляемость нервных центров продемонстрировал Н. Е. Введенский в опыте на препарате лягушки при многократном рефлекторном вызове сокращения икроножной мышцы с помощью раздражения n. tibialis и n. peroneus (рис. 37).
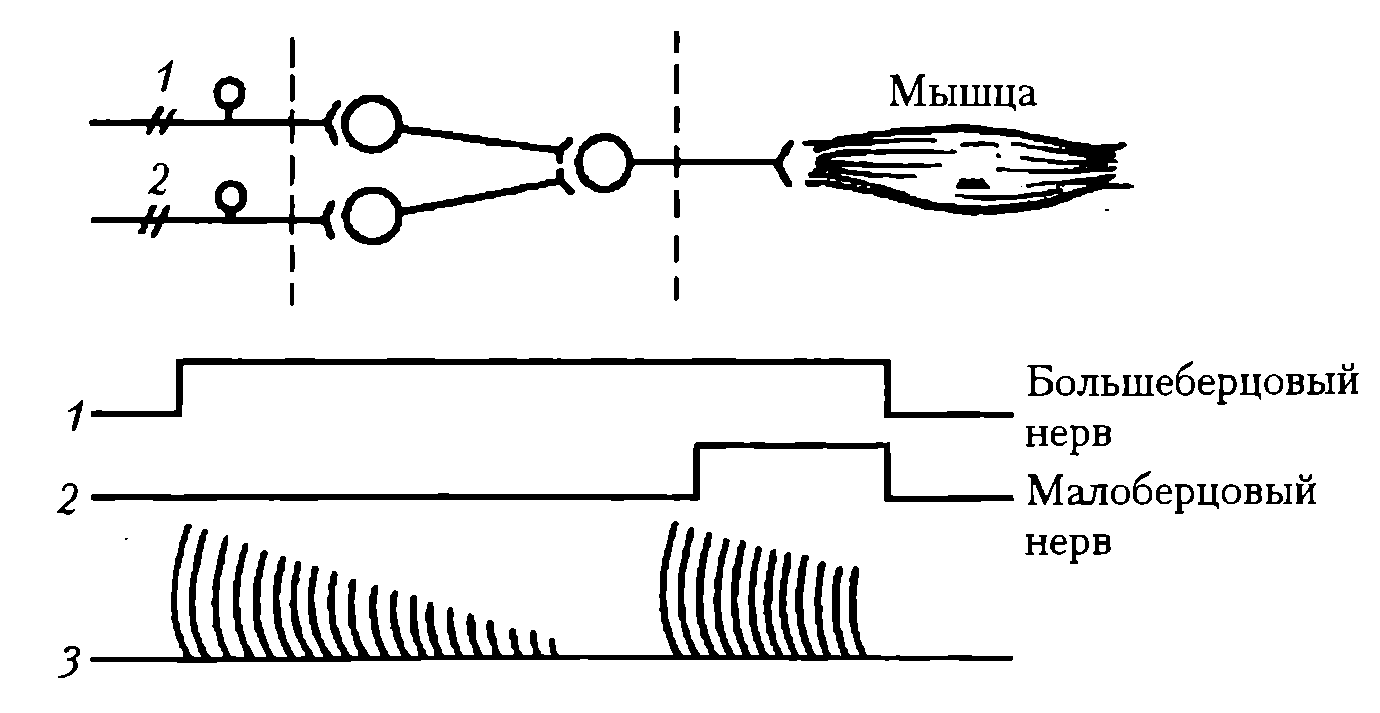
Рис. 37. Схема опыта Н.Е. Введенского, иллюстритрующие локализацию утомления в рефлекторной дуге: 1 – раздражение большеберцового нерва; 2 – раздражение малоберцового нерва; 3 – кривая сокращений полусухожильной мышцы лягушки
В этом случае ритмическое раздражение одного нерва вызывает ритмическое сокращение мышцы, которое постепенно ослабевает вплоть до полного прекращения. Переключение раздражения на другой нерв сразу же вызывает сокращение той же мышцы, что свидетельствует о локалиации утомления не в мышце, а в центральной части рефлекторной дуги (см. рис. 37). При этом развивается ослабление реакции нейронного пула на афферентные импульсы, выражающееся в снижении постсинаптических потенциалов во время длительного раздражения или после него. Оно объясняется расходованием медиатора, снижением чувствительности постсинаптической мембраны к медиатору вследствие накопления метаболитов, закислением среды при длительном проведении возбуждения по одним и тем же нейронным цепям.
Особо важным вариантом пластичности нервных центров является компенсация нарушенных функций после повреждения того или иного центра. Хорошо известны клинические наблюдения за больными, у которых после кровоизлияний в вещество мозга повреждались центры регуляции мышечного тонуса и акта ходьбы. Тем не менее, со временем отмечалось, что парализованная конечность у таких больных постепенно начинает вовлекаться в двигательную активность, при этом нормализуется тонус ее мышц. Нарушенная двигательная функция частично, а иногда и полностью восстанавливается за счет большей активности сохранившихся нейронов и увеличения ветвления их аксона (спраутинг), а также вовлечения в эту функцию других, «рассеянных» нейронов в коре большого мозга с подобными функциями. В случае дополнительного ветвления аксона нейрона возрастает возможность улучшения иннервации мышечных волокон. Этому способствуют регулярные пассивные и активные движения органа, функция которого нарушена. Процессы возбуждения в ЦНС постоянно взаимодействуют с процессами торможения, что обеспечивает согласованную деятельность нервных центров.
ЦНС чувствительна к изменениям внутренней среды, например к изменению газового состава крови, содержания глюкозы в крови, температуры, а также к вводимым с лечебной целью различным фармакологическим препаратам. В первую очередь реагируют синапсы нейронов. Особенно чувствительны нейроны ЦНС к недостаточности глюкозы и кислорода. Если содержание глюкозы в крови в 2 раза ниже нормы, могут возникнуть судороги. Тяжелые последствия для организма вызывает недостаток кислорода в крови. Прекращение кровотока всего лишь на 10с приводит к очевидным нарушениям функций мозга, человек теряет сознание. Прекращение кровотока на 8—12 мин вызывает необратимые нарушения деятельности мозга: погибают многие нейроны, в первую очередь корковые, что ведет к тяжелым последствиям. Это необходимо учитывать во время оперативных вмешательств на сердце, при реанимации пострадавшего.
Дата добавления: 2016-02-16; просмотров: 3304;