Постсинаптическое торможение
Торможение — активный нервный процесс, результатом которого является прекращение или ослабление возбуждения. Торможение всегда возникает как следствие возбуждения. При этом тормозные нейроны активируются медиаторами возбуждающих нейронов. Торможение в ЦНС открыл И.М. Сеченов (1863). В опыте на таламической лягушке он определял латентное время сгибательного рефлекса при погружении задней конечности в слабый раствор серной кислоты. Латентное время рефлекса значительно увеличивается, если на зрительный бугор предварительно положить кристаллик поваренной соли. Ф. Гольц (1870) обнаружил проявления торможения шагательного рефлекса у спинальной собаки при механическом раздражении хвоста. Наличие специальных тормозных структур в продолговатом мозге доказал X. Мегун (1944). В опытах на кошках при изучении разгибательного рефлекса он установил, что раздражение медиальной части РФ продолговатого мозга тормозит рефлекторную активность спинного мозга.
Известно две разновидности торможения: пре- и постсинаптическое.
Открытие постсинаптического торможения. Д. Экклс (1952) при регистрации потенциалов мотонейронов спинного мозга у кошки во время раздражения мышечных афферентов группы 1а зарегистрировал в мотонейронах мышцы антагониста гиперполяризационны мость мотонейрона, что и обусловило название этого потенциала — тормозной постсинаптический потенциал (ТПСП).
На рис. 38 приводится графическое изображение ТПСП на фонепотенциала покоя (ПП) и возбуждающего постсинаптического потенциала (ВПСП).
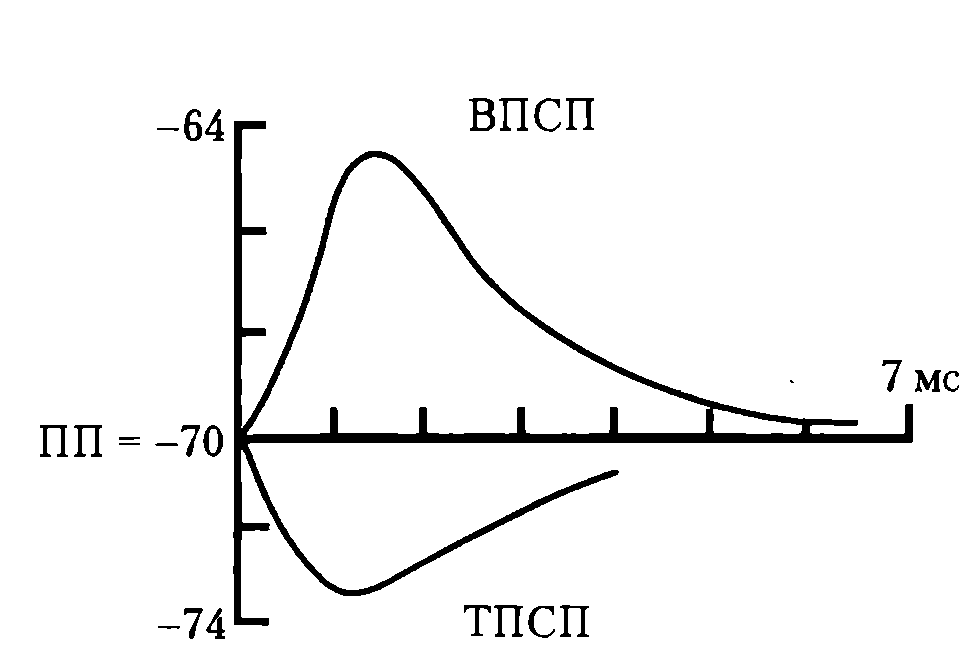
Рис. 38. Возбуждающий и тормозной постсинаптические потенциалы (ВПСП и ТПСП): ПП – потенциал покоя
Амплитуда ТПСП составляет 1-5 мВ; этот потенциал подчиняется закону силы — мощный афферентный залп вызывает возрастание амплитуды ТПСП, способен суммироваться при повторных стимулах.
Механизм постсинаптического торможения. Возбудимость нейрона в процессе развития ТПСП уменьшается потому, что увеличивается пороговый потенциал (ΔV), так как критический уровень деполяризации остается на прежнем уровне, а мембранный потенциал возрастает. ТПСП возникает под влиянием ГАМК и глицина. В спинном мозге глицин выделяется тормозными клетками (клетками Реншоу) в синапсах, образуемых ими на мембране мотонейронов. Действуя на ионотропный рецептор постсинаптической мембраны, глицин увеличивает ее проницаемость для Cl-, при этом последний поступает в клетку согласно концентрационному градиенту, но вопреки электрическому, в результате чего развивается гиперполяризация. В бесхлорной среде тормозная роль глицина не реализуется. Ареактивность нейрона к возбуждающим импульсам является следствием алгебраической суммации ТПСП и ВПСП, в связи с чем деполяризация нейрона не достигает КП.
При действии ГАМК на постсинаптическую мембрану ТПСП развивается в результате входа Cl- в клетку или выхода К+ из клетки, что зависит от вида рецепторов, на которые действует ГАМК.
Имеются ГАМК-рецепторы двух типов: ГАМК1 (ГАМКа) и ГАМК2 (ГАМКв). Активация ГАМК1-рецепторов (они ионотропные) ведёт к непосредственному повышению проницаемости клеточной мембраны для Cl-, который поступает в клетку, как и при действии глицина, согласно концентрационному, но вопреки электрическому, в результате чего развивается гипериоляризация и снижение возбудимости нейрона. Активация ГАМК2-рецепторов (они метаботропные) реализуется с помощью вторых посредников (цАМФ), при этом повышается проницаемость для К+, который выходит из клетки тоже согласно концентрационному градиенту, но вопреки электрическому, в результате чего также развивается гиперполяризация нейрона (ТПСП), что снижает его возбудимость.
Как известно, вышедший из клетки при развитии торможения К+ транспортируется Nа/К-помпой обратно в клетку, что и поддерживает градиент его концентрации. Однако следует признать, что и С1- в процессах торможения транспортируется из клетки специальной хлорной помпой, хотя обычно отмечают, что градиент концентрации С1- поддерживается отрицательным электрическим зарядом внутри клетки — С1- выталкивается отрицательным электрическим зарядом. Поскольку возникновение ТПСП обеспечивается входом С1- в клетку, это доказывает, что концентрационный градиент действует сильнее противоположно направленного электрического градиента. Отсутствие же хлорной помпы привело бы к накоплению С1- в клетке, наступлению равновесия между электрическим и химическим (концентрационным) градиентом для С1- и к нарушению процесса торможения. Но этого не происходит, что и доказывает наличие хлорной помпы.
ТПСП могут возникать вследствие уменьшения проницаемости мембраны и для Na+, что также сопровождаетcя гиперполяризацией клеточной мембраны, особенно если проницаемость для К+ и С1- сохраняется прежней. Такого рода ТПСП были зарегистрированы в нейронах симпатических ганглиев, которые, как известно, локализуются экстраорганно.
Имеется несколько разновидностей постсинаптического торможения.
Возвратное постсинаптическое торможение — тормозные вставочные нейроны действуют на те же нервные клетки, которые их активируют с помощью своих коллатералей. В этом случае развивающееся торможение бывает тем глубже, чем сильнее было предшествующее возбуждение. Типичным примером возвратного постсинаптического торможения является торможение в мотонейронах спинного мозга.
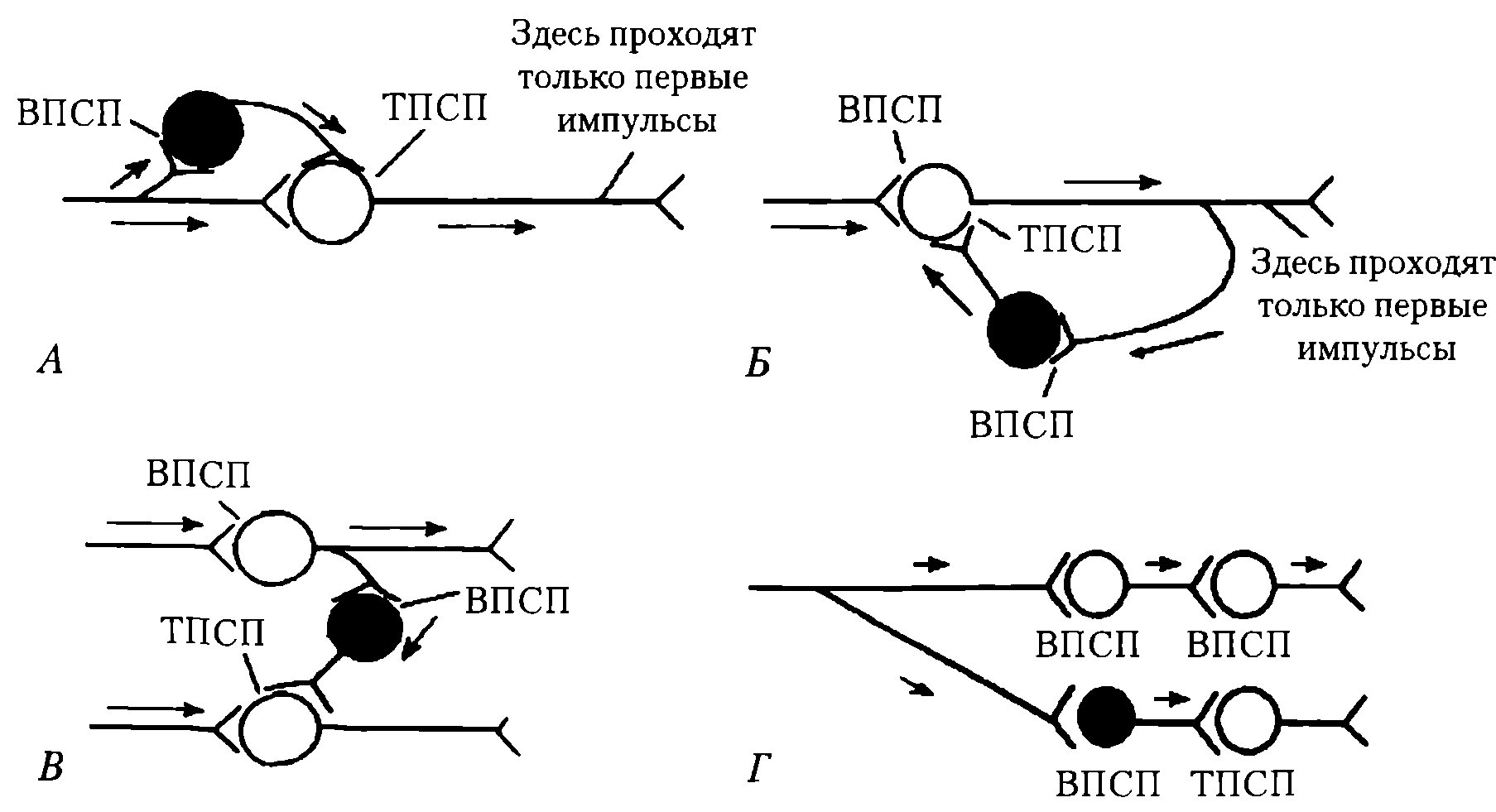
Рис. 39. Разновидности постсинаптического торможения: А – параллельное; Б – возратное; В – латеральное; Г – прямое. Нейроны: светлые – возбуждающие, черные - тормозные
Как видно из рис. 39, А, мотонейроны посылают коллатерали к тормозным вставочным нейронам и возбуждают их с помощью ацетилхолина, действующего на N-холинорецепторы клетки Реншоу.
Аксоны клеток Реншоу, в свою очередь, образуют синапсы на тех же мотонейронах, которые возбуждают тормозную клетку. Такая тормозная цепь называется торможением Реншоу — по имени ученого, который ее открыл. Это торможение в центрах мышц-сгибателей и разгибателей обеспечивает, например, поочередное сокращение и расслабление скелетной мышцы, что необходимо при ходьбе и беге, ритмических движениях верхних конечностей.
Параллельное постсинаптическое торможение выполняет такую же роль — возбуждение блокирует само себя за счет распространения по коллатералям с включением тормозных клеток на своем пути и поступлением импульсов от тормозных клеток к нейрону-мишени (см. рис. 39, Б). В данном случае первый залп импульсов активирует постсинаптический нейрон, а следующий блокируется.
При латеральном постсинаптическом торможении тормозные вставочные нейроны активируются импульсами от возбужденного центра и влияют на соседние клетки с такими же функциями. В результате в соседних клетках развивается очень глубокое торможение. Латеральное торможение может образовать тормозную зону, которая окружает возбужденные нейроны (см. рис. 39, В). Торможение такого типа называется латеральным потому, что образующаяся зона торможения находится сбоку по отношению к возбужденному нейрону и инициируется им.
Прямым постсинаптическим торможением является, например, реципрокное торможение (от лат.—reciprocus взаимный). Так, при раздражении кожных рецепторов возникает защитный сгибательный рефлекс: центр сгибания возбужден, а центр разгибания заторможен. В этом случае возбуждающие импульсы поступают к центру мышцы-сгибателя, а через тормозную клетку — к центру мышцы-антагониста (разгибателю), что предотвращает ее сокращение (см. рис. 39, Г). Если бы возбуждались одновременно центры мышц сгибателей и разгибателей, сгибание конечности в суставе было бы невозможным. При возбуждении центра мышцы-разгибателя угнетается центр мышцы-сгибателя. В реципрокных взаимоотношениях находятся центры дыхания и глотания, вдоха и выдоха.
Дата добавления: 2016-02-16; просмотров: 8399;