Закономерности наследования качественных признаков при аллельном взаимодействии генов
3.1. Закономерности наследования при внутривидовой гибридизации в случае моногибридного скрещивания
Проблему наследственности не удавалось разрешить, пока ее связывали лишь с вопросом сходства родителей и потомства. Грегору Менделю пришла мысль изучить характер наследования тех признаков, которые отличают родителей друг от друга. Он открыл наличие закономерностей, которые имеют большое значение в генетике, а вместе с этим и в селекции.
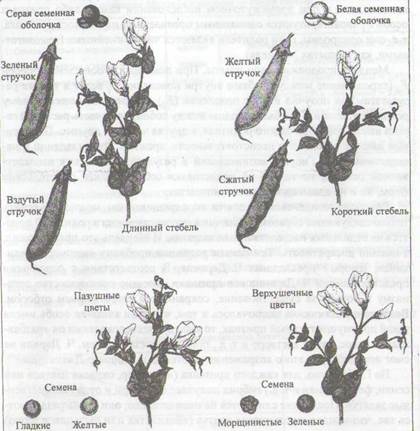
В качестве объекта Мендель выбрал горох — Pisum sativum (самоопыляющееся растение) и именно те формы, которые отличались между собой заметными признаками. Каждая из этих форм (когда возможность оплодотворения другой формой была исключена) в ближайших и последующих поколениях давала потомство, сходное по всем признакам с родителями. Таких признаков родителей было выбрано семь с альтернативным их проявлением (рис. 3.1):
1) форма зрелых семян (круглые, или гладкие, — морщинистые);
2) окраска семядолей (желтые — зеленые);
3) окраска семенной кожуры (серая — белая), которая коррелирует с окраской цветков (пурпурные или красные — белые);
4) форма зрелых бобов (выпуклые — с перехватом);
5) окраска незрелых бобов (зеленые — желтые);
6) расположение цветков (пазушное — верхушечное);
7) высота стеблей (высокие — низкие).
Затем Г. Мендель их скрещивал. Гибрид, полученный в результате скрещивания чистых рас с противоположными признаками, всегда был сходен с одним из родителей. Так, например, по окраске цветков, форме и цвету семян доминировали красные цветки, круглые и желтые семена. Остальные признаки исчезали. Видимый признак (в первом поколении F,) Мендель назвал доминантным, а невидимый, «исчезнувший», — рецессивным.
Полученный им результат, а именно единообразие потомства в Fr поставил под сомнение правильность прежних представлений, согласно которым наследственное вещество обоих родителей смешивается подобно двум жидкостям и потом передается потомству. Позднее путем скрещивания действительно было получено потомство (F;), которое заняло среднее положение между обоими родителями. Например, при скрещивании красноцветковой и белоцветковой форм ночной красавицы — Mirabilis jalapa
38
ДОМИНАНТНЫЕ РЕЦЕССИВНЫЕ
Рис. 3.1. Семь признаков гороха — Pisum sativum, наследование которых изучал Мендель (Ф. Айала, Дж. Кайгер, 1988)
получилось потомство с розовыми цветками. Такой характер появления признаков у гибридов называется промежуточным.
Если материал не изучен в генетическом отношении, нельзя предсказать, каким будет тот или иной признак — доминантным или рецессивным. Полная доминантность или полная рецессивность являются крайними случаями, между которыми существует постепенный переход от ясно выраженного доминирования к среднему состоянию и далее к полной рецессивности. Понятно, что факт полного подавления рецессивного признака доминантным имеет для наследственности и селекции чрезвычайно важное значение. Но говорить на основании исследований Менделя о за-
коне доминирования, как это часто делалось раньше, было бы неправильно.
Лучше говорить об однородности гибридов. Например, как при доминантном, так и при промежуточном наследовании какого-либо признака особи Ft характеризуются одинаковым проявлением изучаемого признака, т. е. они однородны, если родители являются чистолинейными (гомозиготными, как в опытах Менделя).
Мендель продолжил свои опыты. При помощи самоопыления особей F, (скрещивание между особями внутри поколения F. ведет к тем же результатам) он получил второе поколение (F2) и поразился неожиданному результату: особи F2 не были сходны между собой; одна часть растений гороха имела признаки одного родителя, а другая часть — другого. Полученные данные говорили о несостоятельности прежних представлений о наследственности. Гибрид, возникавший в результате смешения наследственной основы, не только не представлял собой среднее родительских форм, но и не давал однородного потомства.
Результат менделевского опыта со скрещиванием можно объяснить только следующим образом: различия в наследственности родителей покоятся на отдельных наследственных задатках. И Мендель это предположил, а именно дискретность. Тем самым разрешив проблему «кошмара Дженкина», который преследовал Ч. Дарвина. В соответствии с основными представлениями Ч. Дарвина все признаки, дающие преимущество организму в борьбе за существование, сохраняются естественным отбором. Возражение Дженкина заключалось в том, что если какая-то особь имела такой преимущественный признак, то в следующем поколении он «разбавлялся» вдвое, затем вчетверо и т. д., пока не исчезал совсем. Ч. Дарвин не смог опровергнуть этого возражения — отсюда и «кошмар Дженкина».
По Г. Менделю, для каждого признака (например, окраска цветков или семян, форма семян и т. д.) гибрид получает от матери и отца наследственные задатки. Последние сливаются не окончательно; они вновь разделяются так, что каждая зародышевая клетка (яйцеклетка или пыльцевое зерно) получает каждый раз задатки только одного представителя пары. Наследственные задатки, как уже отмечалось ранее, названы Иогансеном «генами». Здесь термин «ген» употребляется не как структурная, а как счетная единица, при этом доминантные гены обозначаются прописными, а рецессивные — строчными буквами, например Act, Bb и т. д.
Опять вернемся к опытам Г. Менделя. В поколении F, получились следующие результаты (табл. 3.1).
В этих опытах во втором поколении родительские формы представлены в отношении, близком 3:1. Мендель предположил, что приведенные в таблице числа представляют собой лишь случайные отклонения от действительного отношения 3:1. Одновременно он дал теоретическое объяс-
| Таблица 3.1 | ||
| Распределение особей по разным признакам в поколения F2 в опытах Менделя с горохом — Pisum sativum | ||
| Признаки родителей | F, | | |
| 1. Семена круглые х морщинистые | круглые | 5479 круглые:1850 морщинистые = 2,96:1 |
| 2. Семядоли желтые х зеленые | желтые | 6022желтые:2001 зеленые = 3,01:1 |
| 3. Цветки пурпурные х белые | пурпурные | 705 пурпурные:224 белые = 3,15:1 |
| 4. Бобы выпуклые х с перехватами | выпуклые | 882 выпуклые.299 с перехватами = 2,95:1 |
| 5. Незрелые бобы зеленые х желтые | зеленые | 428 зеленые: 152 желтые Е 2,82:1 |
| 6. Расположение цветков | пазушное | 651 пазушные:207 верхушечные = 3,14:1 |
| 7. Высота стеблей | высокие | 787 высокие:277 низкие = 2,84:1 |
| В среднем | 2,98:1 |
нение (предположение), которое полностью подтвердилось. Вот оно: гибрид образует как женские, так и мужские половые клетки в одинаковом количестве — 50 % яйцеклеток с геном А и 50 % яйцеклеток с геном а. Такое же положение характерно и для мужских гамет. При совершенно случайном оплодотворении половина яйцеклеток каждой категории оплодотворяется одним видом пыльцы, а другая половина — другим, как это показано на следующей схеме, так называемой решетке английского генетика Р. Пеннета (табл. 3.2).
| Комбинация менделевскнх зачатков | Таблица 5.2 | |||
| Пыльца Яйцеклетки ^~"--^. | 50% А | 50% а | ||
| 50% А | 2i% AA | 25 % аА | ||
| 50% а | 25%Ло | 25 % аа |
Таким образом, во втором поколении три различных наследственных типа (АА, Аа, и аа) относятся друг к другу, как 1:2:1. При промежуточном проявлении признака это соотношение сохраняется и в фенотипе. При доминировании получается фенотипическое отношении 3:1, так как особи АА и Аа внешне неотличимы.
Является ли носитель доминирующего признака гомо- или гетерозиготным, т. е. принадлежит ли он к наследственному типу АА или Аа, можно выяснить только по потомству. Опыты Менделя показали, что среди особей с доминантным признаком в среднем только V3 при самоопылении дает константное потомство с доминирующим признаком, в то время
как 2/3 расщепляются подобно первоначальному гибриду. Наоборот, носитель рецессивного признака размножается (при самоопылении или родственном опылении) всегда в чистом виде. Выведенная из численных данных гипотеза, что гибрид, родители которого различаются по одному признаку, образует два типа гамет (50 % А и 50 % а), считается одним из главных достижений Г. Менделя.
Доказательством правильности этого предположения служит опыт с обратным (возвратным, реципрокным) скрещиванием. Если скрещивать особь с доминирующим признаком из поколения F, с рецессивным родителем (при промежуточном проявлении признаков безразлично, какой из родителей будет выбран), то потомство расщепляется в отношении 1:1. Этого и следовало ожидать, если гибрид образует 50 % Л-гамет и 50 % я-гамет. Таким способом можно установить, к какому типу принадлежит носитель доминирующего признака — к типу АА итгАа. Возвратные скрещивания называют еще анализирующими, насыщающими (беккроссами).
3.2. Закономерности наследования при внутривидовой гибридизации в случае дигибридного скрещивания
Итак, при моногибридных скрещиваниях Г. Мендель открыл два правила:
единообразия гибридов в Ft,
расщепления гибридов в F2.
Мендель продолжил свои исследования, проводя дигибридные скрещивания. Он скрещивал горох, имеющий круглые желтые семена, с горохом, у которого семена были зелеными и морщинистыми.
Все растения F, сохраняли единообразие: они имели желтые и круглые семена.
В F2 расщепление носило более сложный характер, чем это наблюдалось при моногибридном скрещивании. Из общего количества (556) полученных в этом опыте семян были:
желтые круглые (гладкие) — 315 шт.,
желтые морщинистые— 101 шт.,
зеленые круглые (гладкие) — 108 шт.,
зеленые морщинистые — 32 шт.,
что соответствует кратному отношению 9:3:3:1.
Сущность явлений при дигибридном скрещивании можно объяснить следующим образом.
В зиготу, из которой развивается гибридное растение Ft, вносится четыре гена: желтой окраски семян (А) и округлой формы их (В) от одного родителя и зеленой окраски семян (а) и морщинистой формы их (Ь) от другого родителя. Такое растение будет дважды гетерозиготным, или дигете-
розиготным. Все возможные сочетания генов дают четыре типа яйцеклеток и четыре типа спермиев: АВ, АЬ, аВ, ab.
 Для расчета сочетаний разных типов и определения результатов расщеплений в F2 строим решетку Р. Пеннета (рис. 3.2).
Для расчета сочетаний разных типов и определения результатов расщеплений в F2 строим решетку Р. Пеннета (рис. 3.2).
Во время мейоза у гибридного растения Ft две материнские хромосомы, несущие неаллельные доминантные гены, и две отцовские хромосомы, несущие неаллельные рецессивные гены, расходятся в дочерние клетки независимо друг от друга и поэтому при случайном соединении гамет во время оплодотворения в F2 образуется девять генотипически разных зигот. Только две из них (ААВВ и aabb) воспроизведут исходные родительские генотипы, остальные же семь будут иметь различные сочетания хромосом с доминантными и рецессивными генами. Этот результат Мендель предвосхитил гипотезой независимого комбинирования генов, которую он вывел из анализа результатов дигибридного скрещивания. А результаты оказались следующими: 1. По фенотипу гибриды F2 образуют четыре класса, которые распределяются в числовом отношении: 9 желтых круглых, 3 желтых морщинистых, 3 зеленых гладких, 1 зеленый морщинистый.
2. Распределение тех же гибридов по генотипу дает 9 классов в следую
щих числовых отношениях:
ААВВ — 1, ААВЬ — 2, ААЪЬ — 1, АаВВ — 2, АаВЬ — 4, АаЬЪ — 2, ааВВ — 1, ааВЪ — 2, aabb — 1.
3. Гены каждой аллельной пары (А-а и В-Ь) распределяются, как и при моногибридном скрещивании, в отношении: 1:2:1 {AAA: %Аа: Ааа и ABB: Ш: Abb).
4. В соответствии с этим и распределение классов по фенотипу по каждой паре аллелей идет в отношении 3:1 (12 желтых:4 зеленых и 12 круг-лых:4 морщинистых).
5. Числовые отношения распределения классов по фенотипу и генотипу при скрещивании организмов, различающихся по двум аллелям, являются результатом произведения числовых отношений по каждой из аллельных пар.
Так, (3:1) х (3:1) = 9:3:3:1 и (1:2:1) х (1:2:1)= 1:2:1:2:4:2:1:2:1 (или
4:2:2:2:2:1:1:1:1).
Это положение верно для любого числа аллелей.
6. Окраска и форма семян у гибридов F2 сочетается не только в тех комбинациях, которые были у родительских форм, но и во всех других возможных комбинациях. Благодаря этому, во втором поколении получаются гибриды, сочетающие признаки обоих родителей (растения с желтыми морщинистыми и зелеными гладкими семенами), т. е. идет новообразование. На этом основана комбинационная селекция. Правильность своих выводов о независимом комбинировании генов при дигибридном скрещивании Мендель проверил путем анализирующего скрещивания гибридных растений в F2, имевших генотип АаВЬ, с отцовским родителем — гомозиготной рецессивной формой по обеим парам генов {aabb). В результате такого скрещивания получилось четыре типа форм: АаВЬ (желтые круглые), Aabb (желтые морщинистые), ааВЬ (зеленые круглые) и aabb (зеленые морщинистые). В каждой из этих групп было одинаковое число особей. Так как во всех четырех скрещиваниях от родительского сорта передавались одинаковые гаметы — ab, то равное число особей во всех четырех группах анализирующего скрещивания является результатом того, что гибриды F, {АаВЬ) образовали яйцеклетки AB, Ab, аВпаЬъ равных количествах, а это возможно только на основе независимого комбинирования генов.
Независимое комбинирование генов и основанное на нем расщепление в F2 в отношении 9:3:3:1 были установлены для большого числа животных и растений, включая и лесные деревья.
наследования признаков при неполном доминировании
 Следует иметь в виду, что расщепление в F2 при дигибридном скрещивании в отношении 9:3:3:1 наблюдается только при полном доминировании по обеим парам аллельных генов. При неполном доминировании, или доминировании по одному признаку, гетерозиготные особи будут отличаться от гомозиготных, и число фенотипических классов в F2 увеличится. Поясним это на примере скрещивания двух форм львиного зёва, имеющих красные нормальные и белые пилорические цветки (рис. 3.3).
Следует иметь в виду, что расщепление в F2 при дигибридном скрещивании в отношении 9:3:3:1 наблюдается только при полном доминировании по обеим парам аллельных генов. При неполном доминировании, или доминировании по одному признаку, гетерозиготные особи будут отличаться от гомозиготных, и число фенотипических классов в F2 увеличится. Поясним это на примере скрещивания двух форм львиного зёва, имеющих красные нормальные и белые пилорические цветки (рис. 3.3).
У этого растения по окраске цветков наблюдается неаллельное доминирование, а нормальная форма цветка доминирует. Гибриды в Ff будут иметь нормальные по форме и промежуточные по окраске (розовые) цветки. В F2 произойдет расщепление в отношении 3 красных нормальных: 6 розовых нормальных: 2 розовых пилорических: 1 красный пилорический: 3 белых нормальных: 1 белый пилорический, т.е. (3:1) х (1:2:1) = = 3:1:6:2:3:1.
Так как при неполном доминировании распределение генов при образовании гамет остается независимым, число и соотношение по генотипу остается таким же, как и при полном доминировании, т. е. 4:2:2:2:2:1:1:Г:1.
Дата добавления: 2016-04-06; просмотров: 1976;