Биофизика нервного импульса
Нормальное функционирование организма невозможно без обмена информацией между клетками. Одним из способов информационного обмена является возможность генерации и восприятия клетками нервного импульса. В организме существуют так называемые возбудимые клетки, к которым относятся мышечные, нервные и секреторные. Эти клетки способны откликаться каким-либо образом на своё возбуждение. Так, мышечные клетки сокращаются, секреторные выделяют биологически активные вещества, а нервные генерируют электрические колебания – нервный импульс.
Потенциал покоя. На мембране клетки всегда существует разность потенциалов, то есть электрический потенциал внутри клетки отличается от наружного. Эта разность потенциалов называется потенциалом покоя. Если наружный потенциал принять равным нулю, то внутренний составит от 50 до 90 мВ, в зависимости от вида клеток. Причиной возникновения потенциала является наличие градиента концентраций ионов K+, Na+ и Cl на клеточной мембране. Допустим, что мембрана проницаема только для ионов калия и непроницаема для остальных, тогда потенциал покоя будет определяться равновесным калиевым потенциалом и описываться уравнением Нернста. Но на самом деле величина потенциала покоя будет всегда несколько ниже равновесного калиевого потенциала, так как внутрь клетки постоянно просачиваются ионы натрия и приносят туда свой положительный заряд. Диффузия отрицательных ионов хлора, наоборот способствуют увеличению потенциала покоя. То есть вклад в формирование потенциала покоя вносят потоки всех ионов, коэффициенты проницаемости которых отличны от нуля. Зависимость потенциала покоя от концентрации и коэффициентов проницаемости описывается уравнением Гольдмана-Ходжкина-Катца: Δφ = RT/F• lg{{PK[K+]0+PNa[Na+]0+PCl[Cl-]i}/{PK[K+]i+PNa[Na+]i+PCl[Cl-]0}}.
В состоянии покоя мембранные проницаемости для ионов К+, Nа+ и Cl- относятся друг к другу как PK : PNa : PCl = 1: 0,04 : 0,45.
Так как в покое проницаемость клетки для ионов калия намного больше её проницаемости для других ионов, то потенциал покоя определяется преимущественно разностью концентраций ионов калия.
Поддержание разности концентраций ионов осуществляется при помощи работы ионных насосов, использующих энергию АТФ. Na+,K+ - насос, кроме того, способствует увеличению трансмембранного потенциала, так как выводит три иона натрия и закачивает в клетку только два иона калия, что увеличивает положительный заряд межклеточной среды, а, следовательно, увеличивает потенциал покоя.
Увеличение трансмембранной разности потенциалов называется гиперполяризацией, уменьшение – деполяризацией. При деполяризации может измениться знак внутриклеточного потенциала. К уменьшению потенциала покоя приводят:
1. нарушение работы АТФ-азы;
2. действие ядов, которые увеличивают проницаемость мембраны для ионов натрия;
3. торможение процессов, обеспечивающих синтез АТФ в клетке.
Потенциал действия. При воздействии на клетку какого-либо раздражителя её трансмембранный потенциал изменяется, возникает так называемый потенциал действия (ПД), или спайк. Причиной такого колебания потенциала покоя является изменение проницаемости мембраны для натрия, что, в свою очередь, вызвано открытием натриевых ионных каналов. При возбуждении проницаемости мембраны для ионов калия и натрия относятся друг к другу как PK : PNa=1:20. В результате поток ионов натрия в клетку начинает превышать поток ионов калия из клетки. Если раньше потенциал на мембране был близок к равновесному калиевому потенциалу, то теперь он стремится к равновесному натриевому, но не достигает его только вследствие того, что проводимости для ионов калия и хлора отличны от нуля. В клетке происходит деполяризация мембраны: отрицательный потенциал клетки приближается к нулю, а потом и вовсе меняет знак на противоположный. Последний процесс называется реверсией мембранного потенциала. Максимальное значение потенциала действия составляет обычно 30-40 мВ. Увеличение натриевой проводимости длится доли миллисекунд. Далее она начинает снижаться, а калиевая – возрастать, в результате чего в клетке восстанавливается потенциал покоя. Этот процесс называется реполяризацией клеточной мембраны.
Для восстановления потенциала покоя клетке требуется некоторое время. В процессе реполяризации сначала происходит быстрое приближение к нормальному значению потенциала покоя, затем скорость изменения потенциала уменьшается, и клетка некоторое время пребывает в деполяризованном состоянии. Это называется следовой деполяризацией. В некоторых клетках, наоборот, возникает следовая гиперполяризация, то есть в процессе реполяризации разность потенциалов начинает превышать обычную. Оба эти отклонения от нормального значения потенциала покоя называются следовыми потенциалами.
Действие раздражителя обычно приводит к локальной деполяризации мембраны. Это вызывает открытие натриевых каналов, чувствительных к изменению потенциала, а, следовательно, - увеличивает натриевую проводимость, что приводит к ещё большей поляризации. Указанный процесс способствует открытию новых натриевых каналов. Существование такой обратной связи обеспечивает регенеративную или самообнавляющуюся деполяризацию клеточной мембраны. После возникновения потенциала действия данный участок мембраны некоторое время находится в невозбудимом – рефрактерном – состоянии, то есть действие раздражителя не вызывает генерацию потенциала действия, так как натриевые каналы некоторое время после активации находятся в закрытом состоянии и не способны открыться в ответ на изменение трансмембранного потенциала.
Длительность потенциала действия отличается для различных клеток и существенно зависит от температуры. При её уменьшении на 100С время существования потенциала действия увеличивается в три раза. При этом длительность реполяризации обычно превышает длительность деполяризации.
Амплитуда потенциала действия не зависит от силы раздражителя, а сам ПД возникает только в том случае, если деполяризация мембраны превысила некоторый пороговый уровень, определяемый свойствами данной клетки. Это явление получило название закона «всё или ничего». Однако если деполяризация составляет 50-70% от уровня пороговой, то в клетке может возникнуть так называемый локальный ответ, амплитуда которого значительно ниже амплитуды потенциала действия. Чем выше уровень подпороговой деполяризации (то есть чем ближе он к пороговой), тем выше амплитуда локального ответа. Отсутствие потенциала действия при подпороговом уровне деполяризации объясняется тем, что в этой ситуации недостаточно увеличивается натриевая проводимость, а, следовательно, нет возможности вызвать регенеративную деполяризацию. Подпороговый уровень деполяризации не вызывает открытия новых натриевых каналов, поэтому натриевая проводимость быстро уменьшается, и в клетке восстанавливается потенциал покоя.
Амплитуда потенциала действия и пороговый уровень деполяризации не являются строго постоянными величинами для данной клетки. Длительная деполяризация приводит к увеличению инактивации натриевых каналов и активации калиевых. В результате амплитуда потенциала действия уменьшается, а пороговый уровень деполяризации увеличивается. Длительная гиперполяризация вызывает увеличение амплитуды ПД и уменьшение порогового уровня деполяризации.
При действии электрического тока на мембрану клетки выяснилось, что возникновение ПД зависит не только от силы тока, но и от времени его действия. Чем выше сила тока, тем меньше времени требуется подавать его на клетку, чтобы возник ПД. Действия больших токов в течение короткого промежутка времени не вызывает нужного уровня деполяризации, так же как и действие в течение длительного времени токов малой силы. Наименьший ток, который способен вызвать потенциал действия, называется реобазой. Соответствующее ему время возбуждения – полезным временем. В биологии и медицине часто используют термин хронаксия, который означает время, в течение которого должен действовать ток в две реобазы, чтобы вызвать возбуждение. Для каждой клетки существует своё значение реобазы и хронаксии, поэтому измерение этих характеристик имеет диагностическое значение.
Распространение возбуждения по нервному волокну. Нервные волокна делятся на миелинизированные и немиелинизированные. Миелинизированное нервное волокно состоит из осевого цилиндра, содержащего аксоплазму, покрытого цитоплазматической мембраной. Вокруг него многократно обертываются шванновские клетки или олигодендроциты, слои которых сливаются и образуют миелиновую оболочку. Через равные промежутки (от 0,2 до 2 мм) эта оболочка прерывается, и мембрана осевого цилиндра остаётся открытой. Такие участки волокна называются перехватами Ранвье. Их длина составляет около 1 мкм. Миелиновая оболочка, состоящая из мембранных липидов и белков, является изолятором нервной клетки, благодаря ей возбуждение может возникнуть только на оголённом участке мембраны аксона.
Рис. 1. Схема строения миелинизированного нервного волокна
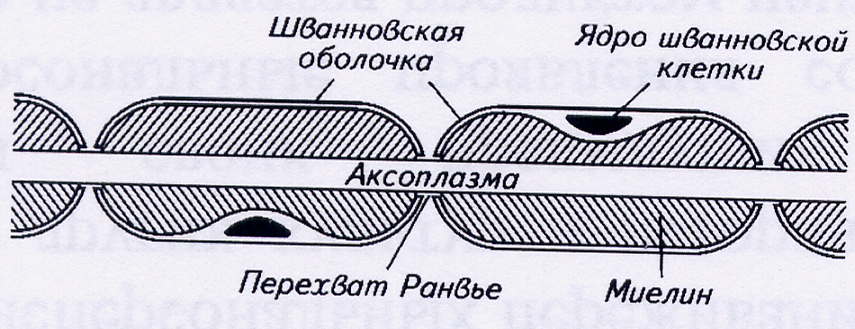
Немиелинизированные нервные волокна не имеют такой плотной жировой оболочки. Шванновская клетка окружает их только один раз.
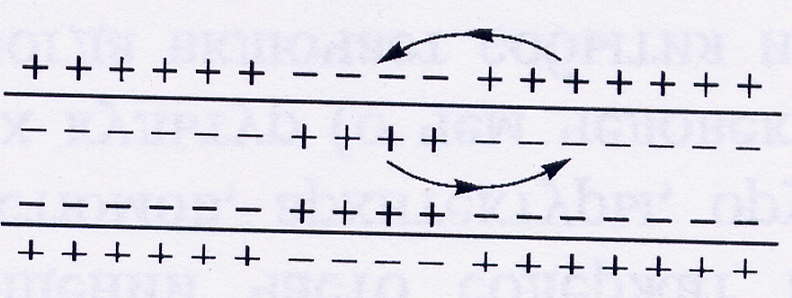
Возбуждение какого-либо участка немиелинизированного нервного волокна приводит к локальной деполяризации мембраны. В то же время остальная часть мембраны сохраняет свою обычную разность потенциалов: наружная среда заряжена положительно, а внутренняя – отрицательно. Между возбужденным и невозбужденным участками возникают местные токи. Это приводит к деполяризации соседнего участка, который, в свою очередь, деполяризует следующий. Такой способ проведения возбуждения называется непрерывным.
Рис.2. Схема механизма проведения нервного импульса по немиелинизированному волокну
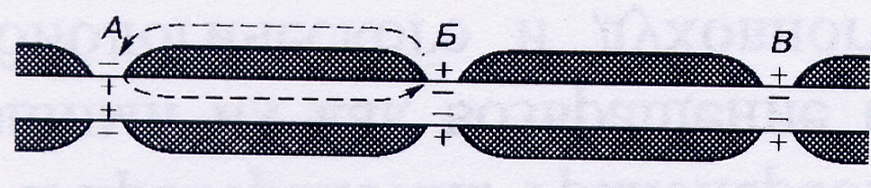
В миелинизированных нервных волокнах непрерывное проведение нервного импульса невозможно. Возбуждение (деполяризация) может возникнуть только в перехватах Ранвье. Деполяризация одного участка вызывает деполяризацию соседнего участка. Далее возбуждение способно перейти только вперед к следующему участку, так первый участок в течение некоторого времени остается рефрактрным. По этой причине нервный импульс по миелинизированному волокну распространяется только в одном направлении. Возникающий потенциал действия в несколько раз превышает порог, необходимый для возникновения возбуждения в следующем перехвате Ранвье, который, таким образом, каждый раз усиливает сигнал, ослабевающий в результате сопротивления межтканевой жидкости и аксоплазмы, и действует подобно ретранслирующему генератору. Механизм распространения возбуждения по миелинизированным волокнам называется сальтаторным.
Рис.3. Схема механизма проведения нервного импульса по миелинизированному волокну
Сальтаторный механизм выгоднее непрерывного, так как позволяет увеличить скорость проведения нервного импульса. Он более экономичен с энергетической точки зрения: деполяризуются только небольшие участки мембраны, возникает меньше потерь ионов, следовательно, клетке приходится расходовать меньше энергии для обеспечения работы Na+K+ - насосов.
Скорость проведения нервного импульса по немиелинизированному волокну пропорциональна квадратному корню из диаметра волокна. Увеличение диаметра способствует увеличению скорости распространения возбуждения.
Для миелинизированных волокон скорость проведения возбуждения зависит от длины межперехватных участков (участков между двумя перехватами Ранвье). В тоже время длина межперехватных участков пропорциональна диаметру волокна. Таким образом, скорость проведения нервного импульса по мякотным волокнам пропорциональна их диаметру.
В зависимости от толщины, а также наличия или отсутствия миелиновой оболочки все нервные волокна делят на три основных типа: А, В, и С.
1. Волокна типа А – это наиболее толстые, хорошо миелинизированные афферентые и эфферентые волокна соматичекой нервной системы. Скорость проведения этих волокон варьирует от 120 м/с до 15 м/с.
2. Волокна типа В - слабомиелинизированные преганглионарные (парасимпатические) волокна вегетативной нервной системы. Скорость проведения составляет 5 – 14 м/с.
3. Волокна типа С – это немиелинизированные в основном постганглионарные (симпатические) волокна вегетативной нервной системы. Скорость проведения от 0,5 до 2,3 м/с.
| <== предыдущая лекция | | | следующая лекция ==> |
| Описание методик проведения спирографии | | | Оперативная хирургия груди |
Дата добавления: 2016-04-02; просмотров: 4422;