Биофизика мембранных процессов
Биофизика мембран изучает структуру биологических мембран, транспорт веществ через них, генерацию и распространение нервного импульса, процессы рецепции и преобразования энергии.
Биологические мембраны выполняют в организме множество функций: они ограничивают клетку и клеточные органеллы, разделяя клетку на отдельные области, что позволяет поддерживать неравновесное состояние организма; формируют межклеточные контакты; обеспечивают механическую защиту клеток, избирательный и регулируемы транспорт веществ, проведение нервного импульса; участвуют в синтезе АТФ; обеспечивают оптимальное расположение в пространстве мембранных белков, которые осуществляют ферментативные реакции; обеспечивают работу рецепторов, иммунной системы.
Структура мембран. В настоящее время общепризнанной является жидкостно-мозаичная модель биологической мембраны, предложенная в 1971 году Николсоном и Сингером. Согласно этой модели структурную основу биологической мембраны составляет фосфолипидный бимолекулярный слой, который выполняет функцию барьера для ионов и водорастворимых молекул и функцию основы, матрицы для мембранных ферментов, рецепторов и других, встроенных в мембраны, белков, гликолипидов и гликопротеидов. В состав липидов плазматических мембран входит много холестерина.
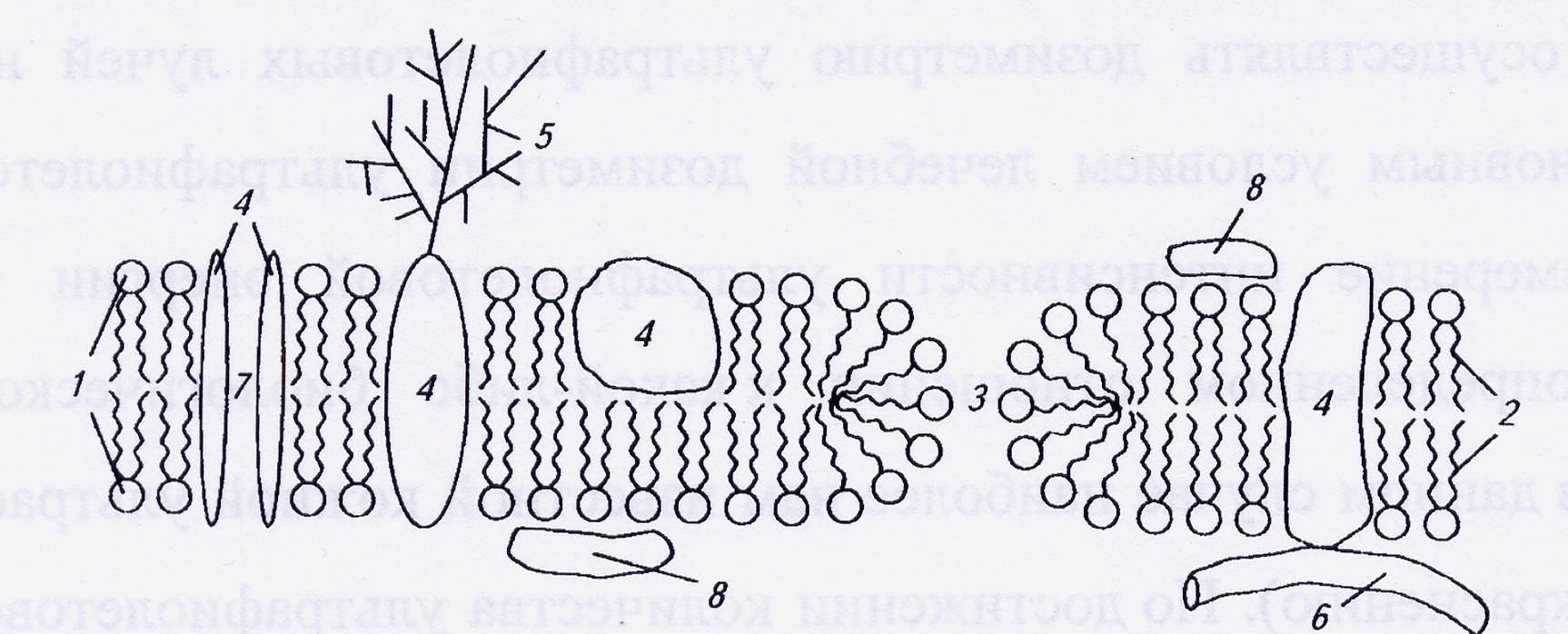
Рис. 1. Схема строения биологической мембраны: 1 – гидрофильные «головки» липидов; 2 – гидрофобные «хвосты» липидов; 3 – гидрофильная липидная пора; 4 – интегральные белки; 5 – углеводная компонента гликолипида; 6 – микротрубочка; 7 – белковый канал; 8 – периферические белки.
Молекулы фосфолипидов обладают свойствами амфифильности, которое заключается в том, что часть молекулы полярна (а потому гидрофильна), а другая – неполярна (и гидрофобна). Это обусловлено особенностями химической структуры фосфолипидов, молекулы которых строятся из глицерина, жирных кислот, фосфорной кислоты и полярного соединения, характерного для каждого класса фосфолипидов (холин, серин, этаноламин и др.). При этом полярную гидрофильную часть называют «головкой», а неполярную гидрофобную – «хвостом». Соотношение длин «головки» и «хвоста» составляет 1:4. Благодаря такому строению фосфолипиды мембран в водной среде стремятся расположиться таким образом, чтобы неполярные «хвосты» контактировали друг с другом, а полярные «головки» - с водой. Поэтому липиды в гидрофильной среде самопроизвольно формируют бислой: внутри оказываются гидрофобные участки, а снаружи – гидрофильные. Такая организация липидных молекул отвечает состоянию с наименьшим значением потенциала Гиббса. Образовавшийся билипидный слой не образует краёв, а стремится замкнуться на себя. Подобное строгое пространственное расположение молекул в мембране во многом определяет её физические свойства. Мембраны имеют высокое удельное электрическое сопротивление, высокую удельную ёмкость и диэлектрическую проницаемость.
Большинство фософолипидов имеют два жирнокислотных остатка. Такие липиды своей формой напоминают сплющенный цилиндр. Если же в липиде одна из двух жирных кислот отсутствует, например, в результате перекисного окисления липидов или действия некоторых фосфолипаз, то молекула напоминает конус. В водном растворе такие липиды образуют мицеллы, а в мембране – гидрофильные поры, через которые способны проходить молекулы воды и ионы, для которых в норме проницаемость мембраны крайне мала. Изменение проницаемости мембраны для веществ может привести к нарушению функционирования и гибели клетки. Так, например, ионизирующее излучение усиливает процессы перекисного окисления липидов, а, следовательно, способствует образованию пор, что является одним из механизмов поражения клетки.
Мембраны содержат большое число различных белков. Их количество так велико, что поверхностное натяжение мембраны ближе к величине поверхностного натяжения на границе раздела «белок-вода», чем «липид-вода». Концентрация мембранных белков зависит от вида клетки. Например, в миелиновой оболочке аксона белков в 2,5 раза меньше, чем липидов, а в эритроцитарной мембране, наоборот, - белков в 2,5 раза больше. Одни белки находятся на поверхности мембраны (периферические), другие частично погружены в липидный слой или пронизывают мембрану насквозь (интегральные белки).
Интегральные белки, так же как и липиды, обладают свойством амфифильности: та часть белка, которая находится в мембране, состоит из гидрофобных аминокислот, скрученных в α-спираль; другая часть, выступающая из мембраны, - гидрофильных аминокислот. Интегральные белки удерживаются в мембране благодаря гидрофобным взаимодействиям. Но иногда их удерживают молекулы цитоскелета – микротрубочки и микрофиламенты. К концу интегрального белка, выступающего во внеклеточную среду, могут быть присоединены углеводы. Эти комплексы называются гликопротеинами. Они часто являются рецепторами, участвующими в иммунных реакциях. Примерами интегральных белков являются белок фоторецепторных мембран – бактериородопсин, пронизывающий мембрану семь раз, белок эритроцитов – гликофорин, к полипептидной цепи которых могут быть присоединены различные олигосахариды; эти гликопротеины определяют группу крови системы АВ0. Многие интегральные белки образуют каналы, через которые могут проходить водорастворимые молекулы и ионы. В этом случае внутренняя часть канала содержит гидрофильные радикалы аминокислот. Интегральные белки связаны с липидами прочными гидрофобными взаимодействиями.
Периферические белки могут находиться как с внутренней, так и с внешней стороны мембраны. Обычно они связаны с липидами электростатическими взаимодействиями, то есть значительно слабее, чем интегральные белки. Поэтому периферические белки легко удаляются с мембраны.
В плазматических мембранах большинства клеток к бислою снаружи примыкает гликокаликс, состоящий из гликолипидов и гликопротеинов. Удельная доля углеводов в биомембранах составляет не более 10-15%.
Толщина биологических мембран составляет 5-10 нм, однако, несмотря на это, их доля в сухой массе клеток превышает 50%. Это объясняется плотной упаковкой компонентов мембран, а также большой суммарной площадью цитоплазматических и внутриклеточных мембран.
Все природные мембраны асимметричны, то есть одинаковые молекулы липидов находятся на наружной и внутренней поверхности в различных концентрациях. Свойство асимметрии присуще и расположению мембранных белков.
Мембранные липиды и белки обладают большой подвижностью, то есть способны диффундировать вследствие теплового движения. Если перемещение их молекул происходит в пределах одного мембранного слоя, то такой процесс называется латеральной диффузией; если же их молекулы перемещаются из одного слоя в другой, то такой процесс называется «флип-флоп» - переход. По сравнению с латеральной диффузией «флип-флоп»-переход происходит гораздо реже. Среднее время между переходами молекулы фосфолипида с одной поверхности мембраны на другую не превышает одного часа. Молекулы белков почти не испытывают таких переходов. Скорость перемещения молекул липидов выше, чем молекул белка. Молекулы перемещаются с помощью поступательного движения, могут совершать и вращательные движения. Подвижность некоторых мембранных белков бывает резко ограничена микротрубочками или микрофиламентами, которые удерживают их в определенном месте на мембране. Большие различия между скоростями латеральной диффузии и «флип-флоп»-переходов очень важны. Столь большая скорость латеральной диффузии обеспечивает высокую скорость химических реакций, происходящих на мембранах, а малая скорость «флип-флоп»-переходов способствует сохранению асимметрии мембран.
В организме позвоночных, а также многих беспозвоночных животных существуют так называемые базальные мембраны. Эти мембраны представляют собой стенки, окружающие группы клеток, часто они расположены между эпителиальной и соединительной тканью, в стенках кровеносных сосудов. Строение базальных мембран принципиально отличается от строения билипидных мембран клеток. Базальные мембраны в основном состоят из гликолипидов и белков. Они обеспечивают механические связи между клетками, играют роль фильтра, особенно в стенках кровеносных сосудов, где они регулируют проникновение веществ из крови в ткань. Система базальных мембран капилляров мозга называется гематоэнцефалическим барьером. Благодаря нему в мозг не проникают многие вещества.
Фазовые переходы в мембранах. При нормальных физиологических условиях мембраны находятся в жидком состоянии, однако, в отличие от жидкостей, они имеют строгую упорядоченную структуру, поэтому это состояние мембран называется жидкокристаллическим. По данным физических методов биологические мембраны способны испытывать фазовый переход первого рода. С понижением температуры мембраны переходят из жидкокристаллического состояния (золь) в твердокристаллическое (гель). При таком переходе сохраняется общая структура мембраны, но порядок в системе еще более возрастает: «хвосты» липидов вытянуты строго параллельно друг другу, их колебания ограничены. Если в золь-состоянии площадь мембраны, приходящаяся на одну молекулу липида, составляет 0,58 нм2, то в гель-состоянии эта величина уменьшается до 0,48 нм2. Толщина мембраны при переходе в гель-состояние увеличивается, но за счет уменьшения площади объём мембраны в целом уменьшается.
Подвижность липидных молекул в обоих фазовых состояниях существенно отличается. В гель-состоянии липиды способны совершать только совместные колебания или вращательные движения. В жидком состоянии липидные «хвосты» имеют большую свободу, особенно велика их подвижность в середине мембраны.
Молекулы липидов жидкой мембраны могут находиться в в двух конформациях: в полностью транс- конформации и в гош-транс-гош-конформации. В гель-состоянии реализуется только транс-конформация. Длина молекулы в гош-конформации меньше, чем в транс-, а ширина, наоборот, больше. Этим объясняется уменьшение толщины и увеличение площади мембраны, приходящейся на один липид, при переходе из гель- в золь-состояние. Способность липидов жидкокристаллической мембраны переходить в гош-конформации способствует повышению её проницаемости для некоторых гидрофильных низкомолекулярных соединений. Если два соседних участка рядом расположенных молекул находятся в гош-конформации, то образуется так называемый кинк. Кинки способны перемещаться вдоль липидных «хвостов». Если небольшая молекула, окажется между двумя кинками, то она сможет пересечь гидрофобный слой.
В последнее время доказано, что при фазовых переходах из золь-состояния в гель- и обратно в мембранах образуются поры диаметром 2-6 нм. То есть при фазовом переходе увеличивается проницаемость мембраны для ионов и низкомолекулярных соединений. Так, проведение нервного импульса определяется изменением проницаемости мембраны для некоторых ионов. Существует предположение, что механизмы терморецепции, также определяются фазовым переходом в мембранах рецепторов. И ещё, так как фазовый переход может иметь и химическую природу (например, липиды мембраны могут перейти в гель-состояние вследствие увеличения концентрации ионов кальция в межклеточной жидкости), то, возможно, эти процессы определяют и хеморецепцию.
Фазовый переход в мембранах происходит не мгновенно, а на протяжении некоторого температурного интервала. Температурой фазового перехода называется та температура, при которой одна половина мембранных липидов находится в золь-состоянии, а другая половина в гель-состоянии. Температура фазового перехода зависит от липидного состава мембран: чем больше в «хвостах» липидов двойных связей (а их число в одном «хвосте» может колебаться от 0 до 4), то есть чем больше степень ненасыщенности жирнокислотных остатков, тем ниже температура фазового перехода. Для мембран, состоящих из насыщенных липидов, эта величина составляет +60  С, а для мембран, состоящих из ненасыщенных липидов, - снижается до -20 С. Такая разница в температурах плавления объясняется тем, что ненасыщенные жирные кислоты, находящиеся в цис-конформации, способствуют удалению друг от друга молекул липидов, и, следовательно, уменьшают силы сцепления между ними. Чем слабее связь между молекулами, тем ниже температура перехода мембран в жидкую фазу.
С, а для мембран, состоящих из ненасыщенных липидов, - снижается до -20 С. Такая разница в температурах плавления объясняется тем, что ненасыщенные жирные кислоты, находящиеся в цис-конформации, способствуют удалению друг от друга молекул липидов, и, следовательно, уменьшают силы сцепления между ними. Чем слабее связь между молекулами, тем ниже температура перехода мембран в жидкую фазу.
Живые организмы, обитающие в разных климатических условиях, имеют различное соотношение насыщенных и ненасыщенных связей в молекулах липидов, что обеспечивает им приспособление к холоду или жаре в зависимости от среды обитания. Более того, это соотношение меняется для разных участков тела одного и того же организма. Например, температура ноги возле копыта полярного оленя может составлять -20 С, а температура ноги возле туловища достигает +30 С. Однако клеточные мембраны не претерпевают фазовый переход за счет того, что мембраны клеток возле копыта содержат больше ненасыщенных липидов, а возле туловища – больше насыщенных. Известно, что животные жиры при комнатной температуре обычно находятся в твердом состоянии, а растительные, наоборот, - в жидком. Это объясняется тем, что в животных клетках меньше ненасыщенных липидов, чем в растительных.
Фазовые переходы мембранных липидов носят кооперативный характер, то есть чем большее количество липидов совершило фазовый переход, тем легче это будет сделать остальным. В идеале, если на каком-либо участке одна молекула липида перешла из твердой фазы в жидкокристаллическую, то за этим сразу же последует фазовый переход всех липидов этого участка (закон «всё или ничего»). Такой участок называется кооперативной единицей, а число молекул, входящих в него, - размером кооперативной единицы n. Для липидных мембран n может достигать нескольких десятков. Величина nтем больше, чем более однороден химический состав мембран. Для уменьшения однообразия липидного состава мембраны животных клеток содержат большое количество холестерина. Например, в эритроцитах млекопитающих его удельная доля от всех липидов составляет 40 - 60%. Такое количество холестерина значительно уменьшает размер кооперативной единицы и снижает различия между твердой и жидкой фазами.
Транспорт веществ через биологические мембраны. Большинство процессов жизнедеятельности, таких, как всасывание, выделение, проведение нервного импульса, мышечное сокращение, синтез АТФ, поддержание постоянства ионного состава и содержания воды связано с переносом веществ через мембраны. Этот процесс в биологических системах получил название транспорта.
Если перенос вещества происходит с уменьшением электрохимического потенциала, то есть не требует затрат энергии, то такой транспорт называется пассивным. Его разновидностями являются диффузия(перемещение веществ в сторону меньшей концентрации) и фильтрация(просачивание веществ поры в сторону меньших значений давления). С помощью диффузии в клетку проникают растворенные молекулы кислорода и углекислого газа, а также яды и лекарственные препараты. Примером фильтрации в организме является перенос воды через стенки кровеносных сосудов, выдавливание плазмы крови в почечные канальцы.
Транспорт веществ через липидный бислой с помощью простой диффузии совершается с малой скоростью, особенно в случае заряженных частиц, и почти не контролируется. Поэтому в процессе эволюции для некоторых веществ появились специфические мембранные каналы и мембранные переносчики, которые способствуют повышению скорости переноса и, кроме того, осуществляют селективный транспорт. Пассивный транспорт веществ с помощью переносчиков называется облегченной диффузией.
Иногда требуется перенести вещество из области с меньшим значением электрохимического потенциала в область с большим его значением. Этот процесс не может протекать самопроизвольно и требует затрат энергии. Такой вид транспорта называется активным. Например, в сторону увеличения электрохимического потенциала осуществляется трансмембранный перенос натрия. Если энергия, необходимая для осуществления активного транспорта, берется за счет гидролиза АТФ или окислительно-восстановительных реакций, то такой транспорт называется первично-активным; если – за счет градиента концентраций других ионов, то – вторично-активным или сопряженным.
Через мембрану могут переноситься не только отдельные молекулы, но и твердые тела (фагоцитоз), растворы (пиноцитоз). Если вещество транспортируется внутрь клетки, то такой вид транспорта называется эндоцитозом, если наружу, то – экзоцитозом. В первом случае на наружной стороне мембраны образуется впячивание, которое постепенно превращается в пузырек. Пузырек отрывается от мембраны внутри клетки. Такой пузырек содержит в себе транспортируемое вещество, окруженное билипидной оболочкой (везикулой). В дальнейшем везикула сливается с какой-нибудь клеточной органеллой и выпускает в неё своё содержимое. В случае экзоцитоза процесс происходит в обратной последовательности: везикула подходит к мембране с внутренней стороны клетки, сливается с ней и выбрасывает своё содержимое в межклеточное пространство.
Пассивный транспорт нейтральных частиц. При наличии градиента концентраций вещества совершается диффузия или пассивный транспорт указанного вещества из области с большей концентрацией в область с меньшей концентрацией. Это явление происходит самопроизвольно (без затрат энергии) до тех пор, пока концентрации не выровняются, и суммарный поток вещества не обратится в ноль (в случае живых клеток такое выравнивание может и не наступить, если вещества непрерывно синтезируются или, наоборот, затрачиваются в ходе химической реакции).
Различают несколько типов пассивного переноса веществ через мембраны: простая диффузия, перенос через каналы и транспорт с помощью переносчиков за счет диффузии переносчика вместе с веществом в мембране (подвижный переносчик) или эстафетной передачи вещества от одной молекулы переносчика к другой (молекулы переносчика образуют временную цепочку поперек мембраны). Все механизмы переноса веществ делят на две группы:
· Такие, при которых каждая молекула переносится независимо от других и эффекты концентрационного насыщения отсутствуют;
· Такие, в которых перенос осуществляется после связывания транспортируемой молекулы переносчиком; по мере заполнения свободных переносчиков наблюдается эффект концентрационного насыщения скорости переноса.
Пассивный перенос вещества вдоль оси х описывается уравнением Фика: Φ = –Ddc/dx, где Φ – поток вещества; D – коэффициент диффузии; dc/dx – градиент концентрации c в направлении x. Знак «-« означает, что поток направлен в сторону меньших значений концентрации, то есть приводит к уменьшению величины концентрационного градиента. Для расчетного описания переноса веществ через биологическую мембрану пользуются з а к о н о м Ф и к а д л я п а с с и в н о г о т р а н с п о р т а в е щ е с т в ч е р е з м е м б р а н у: Φ = –DK/l •(cвн-cвв) = –P(cвн-cвв), где Φ – плотность потока; D – коэффициент диффузии; К – коэффициент распределения вещества между мембраной и окружающей водной фазой; l – толщина мембраны; cвв – концентрация частиц внутри клетки; свн – концентрация частиц снаружи клетки; P – коэффициент проницаемости (см. тему 2). Иными словами, величина потока и скорость транспорта веществ через мембрану прямо пропорциональны коэффициенту распределения, который количественно отражает степень липофильности вещества. Чем больше значение коэффициента распределения, тем лучше вещество растворяется в мембране и с тем большей скоростью переносится через неё. Если рассматривать пассивный переход с позиций превращения энергии, то поток, проходящий через биологическую мембрану, равен: Φ = – uc (dG/dx), где u = D/RT - коэффициент пропорциональности, который зависит от скорости диффузии молекул и называется подвижностью. Таким образом, поток пропорционален концентрации вещества и градиенту термодинамического потенциала в направлении тока.
Пассивный транспорт ионов.
1. В том случае, когда изменяется концентрация вещества и электрический потенциал, уравнения для потоков незаряженных и заряженных частиц сходны: Φ = – uc (dG/dx).
2. Если изменяется еще и стандартный электрохимический потенциал, то уравнение для потока вещества описывается уравнением Теорелла: Φ = – cu • (dμ/dx), где μ – электрохимический потенциал, который зависит от природы вещества и природы растворителя.
3. Если во всей рассматриваемой области диффузии отсутствуют химические превращения вещества и растворитель одинаков, то в этом случае уравнение Теорелла сводится к электродиффузному уравнению Нернста-Планка: Φ = –uRT (dc/dx) –cuz• Fdφ/dx. Это уравнениеописывает пассивный перенос частиц в условиях существования градиентов концентрации вещества и электрического потенциала в растворе или в однородной незаряженной мембране.
4. Если известны концентрации ионов по обе стороны мембраны, трансмембранная разность потенциалов и коэффициент проницаемости, то величину пассивного потока ионов описывают с помощью уравнения Гольдмана: Φ ={zFΔφΡ/RT} • {[c2exp(zFΔφ/RT) -c1]/[1-exp(zFΔφ/RT]}.
5. Если потоки направлены противоположно, то их отношение описывается уравнением Уссинга-Теорелла: Φ2/Φ1 = c1/c2exp[- zFΔφ/RT].
Вывод: пассивный транспорт ионов может происходить только в случае наличия на мембране градиентов концентрации или электрического потенциала и описывается в общем случае уравнениями Нернста-Планка, Гольдмана или Уссинга-Теорелла. Невыполнение этих уравнений свидетельствует о том, что перенос ионов осуществляется активно или через мембранные каналы, где невозможно существование независимых ионных потоков.
Уравнение Нернста. В живом организме по обе стороны любой биологической мембраны находятся ионные растворы, причем концентрации одного и того же иона по обе стороны часто отличаются. Одной из причин установления и поддержания этих градиентов концентраций является различная проницаемость мембраны для тех или иных ионов. В этом случае градиент концентрации одного вещества может поддерживаться за счет наличия градиента концентрации другого вещества и/или разности потенциалов по обе стороны мембраны (так называемой трансмембранной разности потенциалов). Условием установления равновесия между двумя растворами, разделенными полупроницаемой мембраной, с различными концентрациями одинаковых ионов является равенство электрохимических потенциалов по одну и другую сторону мембраны. Значение разности электрических потенциалов, которая устанавливается на мембране при наличии градиента концентраций, рассчитывается с помощью уравнения Нернста: Δφ = φi - φ0 = (RT/zF)•lg (c0/ci).
Равновесие Доннана. В клетке кроме малых ионов находятся еще и заряженные макромолекулы (белки, нуклеиновые кислоты). Как правило, эти макромолекулы заряжены отрицательно. Мембрана проницаема для малых ионов и непроницаема для макромолекул. Для каждого из растворов, находящихся по обе стороны мембраны, необходимо выполнение условия электронейтральности, то есть сумма положительных и отрицательных зарядов всех ионов должна равняться нулю. Для компенсации внутри клетки отрицательных зарядов макромолекул часть отрицательных малых ионов выходит из клетки, а часть положительных зарядов проходит в клетку из межклеточной среды. То есть на мембране возникает так называемая доннановская разность потенциалов, которая составляет φi - φ0 = -RT/F•[R-]i /2c.
Ионный транспорт через каналы. Механизм обычной электродиффузии в живой клетке обеспечивает проницаемость мембран для кислорода и углекислого газа. Этот процесс происходит слишком медленно и плохо контролируется, поэтому клетка не может его использовать для переноса питательных веществ и необходимых для жизнедеятельности ионов. Так, из каждых 1061 ионов, находящихся в водном растворе, только один находится в липидной фазе мембраны. Скорость переноса ионов значительно возрастает, если в мембране существуют ионные каналы. Наиболее распространены каналы для ионов калия, натрия, кальция. Ионные мембранные каналы образованы интегральными белками. Такой канал может быть либо открыт, либо закрыт для транспорта ионов. Эти два состояния реализуются изменением конформации каналообразующих белков, что может быть вызвано изменением условий в клетке и во внеклеточной среде, например, изменением мембранного потенциала.
Так как обычно с помощью каналов транспортируются гидрофильные вещества, то во внутренней полости их имеется большое число гидрофильных химических групп. Каждый канал неоднороден по строению: вдоль его внутренней полости располагаются различные химические группы, сродство ионов к которым неодинаково. Канал может иметь один или несколько ионных центров связывания. Эти центры представляют собой заряженные группы. Когда ион попадает в канал, он связывается с этими группами и таким образом попадает в потенциальную яму. Для того чтобы попасть в другую потенциальную яму, иону требуется преодолеть некоторый потенциальный барьер. Таких барьеров вдоль длины канала может быть несколько, причем высота их, обычно, неодинакова и может изменяться в зависимости от наличия или отсутствия ионов в канале, или изменения трансмембранной разности потенциалов. Так как в канале находятся заряженные группы, то изменение мембранного потенциала является нелинейным. Кроме этого, наличие заряженных групп может привести к непостоянству значений коэффициента распределения по длине канала.
Многие мембранные каналы настолько узки, что ионы не могут в них двигаться в различных направлениях независимо друг от друга: если в какой-либо потенциальной яме уже есть ион, то другой не может в неё попасть. Это относится, например, к калиевым каналам. Встречные потоки ионов натрия через натриевые каналы можно считать независимыми, но если через эти каналы движутся ионы калия, то принцип независимости уже не выполняется. Для многих каналов, в зависимости от их строения, при высоких концентрациях ионов в окружающей среде наблюдается а) э ф ф е к т н а с ы щ е н и я: при увеличении концентрации ионов скорость их переноса повышается, но только до определенного уровня, выше которого скорость не изменяется; б) б л о к и р о в к а: при концентрациях ионов выше некоторого значения скорость переноса начинает снижаться.
При переходе иона из окружающей среды в канал свободная энергия уменьшается. При этом величина потенциального барьера ниже, чем при переходе иона непосредственно через липидный бислой мембраны.
Ионные каналы обладают селективностью, то есть имеют различные величины проводимости для различных ионов. Каждый канал транспортирует преимущественно ионы одного вида, например, натриевый канал – ионы натрия, калиевый – ионы калия. Через катионные каналы не способны проходить анионы и наоборот. Но для ионов того же знака, что и основной ион, канал уже не обладает такой абсолютной селективностью. Проводимость канала для других ионов своего знака ниже, чем для собственного, но отлична от нуля. Например, через специфические натриевые каналы способны транспортироваться также и ионы калия, но в 20 раз медленнее, чем ионы натрия.
Согласно современным представлениям, селективность канала, прежде всего, определяется радиусом транспортируемого иона. Ионы, радиус которых превышает радиус канала, не способны проходить через канал вследствие такого стерического несоответствия. Ионы, обладающие меньшим радиусом, по сравнению с радиусом канала, также не всегда могут пройти через канал. Чем меньше радиус иона по сравнению с радиусом канала, тем меньше вероятность прохождения иона через этот канал. Этот эффект объясняется тем, что ионы в водном растворе всегда находятся в гидратированном состоянии. Перенос же через канал требует сброса нескольких гидратных оболочек, в результате которого ион остается лишь в одной гидратной «одежке». Сброшенная гидратная оболочка заменяется другими полярными соединениями, в роли которых выступают фиксированные дипольные группы канала. Размер иона должен быть таковым, чтобы он мог взаимодействовать с оптимальным числом этих полярных групп. Например, ион калия в своем селективном канале взаимодействует со всеми пятью атомами кислорода, а меньший по размеру ион натрия может связаться в калиевом канале только с двумя атомами кислорода. Следовательно, прочность связи канала с ионом натрия будет минимальна. А если связи иона с дипольными группами нет совсем, то и пройти он через этот канал не может. Следовательно, лучше всего проходит через канал тот ион, который имеет радиус (вместе с одинарной гидратной «одежкой») равный радиусу канала.
Селективность каналов определяется не только зависимостью проводимости канала от радиуса транспортируемого иона, но и химическим строением иона. Например, через натриевые каналы переносятся ионы гидроксиламмония и не переносятся ионы метиламмония, хотя их радиусы равны радиусу канала. Это объясняется тем, что гидроксильная группа гидроксиламмония способна образовывать водородные связи с атомами кислорода, выстилающими внутренний диаметр канала, а метильная группа метиламмония с ними не взаимодействует.
Каналы могут блокироваться некоторыми соединениями, которые связываются с селективными фильтрами или с входом в канал, закупоривая его. Так как молекула-блокатор должна стерически соответствовать участку блокируемого канала, то для каналов определённого вида существуют специфические блокаторы. Например, молекулы тетродоксина блокируют натриевые каналы, а молекулы тетраэтиламмония – калиевые.
Пассивный транспорт веществ с помощью переносчиков. Для некоторых веществ существуют специфические переносчики белковой природы, которые понижают энергию активации, необходимую для переноса вещества через липидный бислой. Переносчики осуществляют так называемую облегченную диффузию. Общий поток веществ через мембрану при участии переносчиков состоит из двух компонент: потока, вызванного простой диффузией, и потока, вызванного облегченной диффузией. Поэтому в данном случае вещества транспортируются с большей скоростью, чем при простой диффузии. Переносчики, осуществляющие пассивный транспорт, переносят вещества из области с большей концентрацией в область с меньшей концентрацией и не требуют затрат энергии.
Переносчики обеспечивают транспорт веществ, которые сами по себе проникают через мембрану крайне медленно из-за высокой гидрофильности. Существуют переносчики для некоторых аминокислот, углеводов, пуриновых и пиримидиновых оснований, нуклеозидов. Таким образом, например, происходит всасывание многих пищевых продуктов в кишечнике, процессы реабсорбции в почках.
Отличительные признаки транспорта веществ с помощью переносчиков:
1. Переносчики обладают высокой химической специфичностью, то есть способны переносить только определённые субстраты (некоторые переносчики даже способны различать L- и D–изомеры одного и того же вещества).
2. Скорость переноса возрастает пропорционально концентрации переносимого вещества, но лишь до некоторой определённой величины, то есть наблюдается эффект насыщения переносчика. Но в реальной клетке часто бывает так, что даже при полном насыщении переносчика при увеличении разности концентраций скорость переноса всё равно будет возрастать за счет простой диффузии.
3. Переносчики способны конкурентно ингибироваться веществами, структура которых сходна со структурой субстрата.
Одним из наиболее изученных случаев пассивного транспорта веществ с помощью переносчиков является перенос глюкозы в эритроциты. Переносчик D-глюкозы способен транспортировать также вещества, структурно напоминающие субстрат, например, D-маннозу, D-ксилозу, D-арабинозу и D-галактозу, но не соответствующие L-сахара. Перечисленные D-изомеры углеводов вызывают конкурентное ингибирование переносчика D-глюкозы, который вынужден переносить эти вещества вместо своего собственного субстрата. Для того, чтобы специфический переносчик сахара перенёс другое вещество, последнее должно иметь в своей структуре шестиуглеродный пиранозный скелет.
Явление насыщения переносчика при высоких концентрациях транспортируемого вещества можно увидеть на примере реабсорбции глюкозы в почечных канальцах. Плазма крови выдавливается под действием гидростатического давления в почечный клубочек, а затем реабсорбируется обратно за исключением продуктов обмена и избыточных количеств воды и электролитов. Если концентрация глюкозы в плазме крови не превышает так называемый почечный порог, величина которого составляет 11 ммоль/л, то глюкоза реабсорбируется полностью. В противном случае наблюдается явление насыщения системы переноса глюкозы и тогда её остаток выделяется с мочой. Почечный порог реабсорбции существует и для некоторых других веществ, например, фосфата, сульфата, аминокислот. Явление насыщения переносчиков иногда может способствовать поддержанию постоянной концентрации данного вещества в организме. В случае глюкозы почечный порог вдвое выше её нормальной концентрации в плазме крови, поэтому почки не могут принимать участия в регуляции содержания глюкозы в крови. Для фосфата же нормальная концентрация в крови почти совпадает с величиной почечного порога, что приводит к выведению излишнего его количества с мочой и поддержанию его концентрации на постоянном уровне.
Индуцированный ионный транспорт. Транспорт ионов с помощью переносчиков называется индуцированным ионным транспортом, а сам переносчик – ионофором. Структура ионофора такова, что снаружи он покрыт неполярными группами, благодаря чему ему выгодно находиться в гидрофобной части мембраны. Внутри ионофора имеется полость, которая покрыта полярными группами, к которым и присоединяются ионы. Многие ионофоры выделены из микроорганизмов, другие имеют синтетическое происхождение. Известны две разновидности ионных переносчиков. Так называемые подвижные переносчики связываются с ионом, диффундируют с ним через мембрану, высвобождают его по другую сторону и возвращаются обратно. Другие переносчики остаются неподвижными относительно мембраны, они формируют мембранные каналы, которые обычно не пронизывают мембрану насквозь. Ионы, пересекая мембрану, вынуждены переходить по цепочке из одной молекулы-переносчика в другую, поэтому такой перенос называется эстафетным.
Рис. 2. Схема пассивного и активного транспорта.
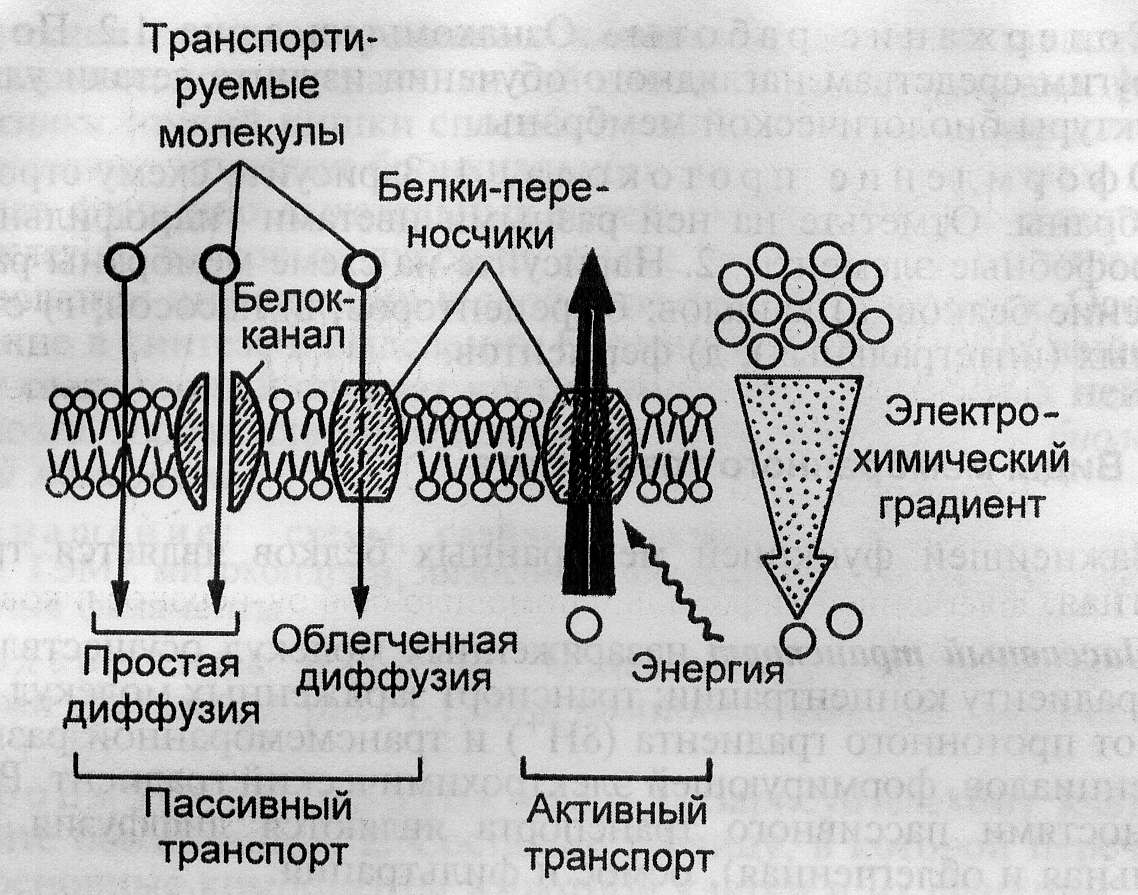
Активный транспорт. В организме часто наблюдается транспорт нейтральных частиц или ионов, в процессе которого электрохимический потенциал возрастает. Например, концентрация ионов калия в клетке намного выше, чем в межклеточной среде, а натрия, наоборот, - ниже. Высокий градиент концентрации калия поддерживается градиентом электрического потенциала, направленным в противоположную сторону, и подчиняется условию равновесия, определяемым уравнением Нернста. Направление градиента электрического потенциала таково, что ионам натрия также гораздо выгоднее находиться в клетке, чем во внеклеточной среде. Самопроизвольный вынос из клетки ионов натрия невозможен и осуществляется только за счет притока энергии. Известны системы активного транспорта дл ионов калия, натрия, кальция и водорода.
Поддержание градиента концентраций ионов натрия и калия обеспечивается работой натрий-калиевого насоса. Он представляет собой мембранные белки, которые работают как ферменты аденозинтрифосфатазы (АТФазы). Задачей этих ферментов является расщепление АТФ на АДФ и неорганический фосфат. Процесс распада сопровождается выделением энергии, которая расходуется на транспорт ионов в сторону увеличения электрохимического потенциала. Расщепление АТФ стимулируется ионами натрия и калия и зависит от наличия магния. Активный транспорт возможен только за счет сопряжения транспорта какого-либо вещества с реакцией гидролиза АТФ.
Механизм сопряжения окончательно не выяснен. Вероятнее всего, энергия АТФ расходуется на изменение конформации транспортного белка, что изменяет его сродство (константу связывания) к тем или иным ионам. Транспорт всегда осуществляется в ту сторону, где сродство ниже. В клетке константа связывания переносчика с Na+ значительно выше, чем с K+. Поэтому ионы натрия в клетке связываются с белком и транспортируются во внеклеточную среду. По другую сторону мембраны конформация белка меняется таким образом, что константа связывания с Na+ уменьшается, а с K+ – увеличивается. Структура ионсвязывающего участка белка в этом случае такова, что к нему могут присоединяться уже не три, а два иона калия, которые и переносятся в клетку.
Согласно современным представлениям, процесс активного транспорта Na+ и K+ происходит в следующие семь этапов.
1. В присутствии Mg2+ на внутренней стороне мембраны образуется комплекс фермента АТФазы с АТФ.
2. Присоединение АТФ изменяет конформацию фермента таким образом, что к образовавшемуся комплексу присоединяются три иона натрия.
3. Происходит фосфорилирование Na+,K+ - АТФазы и отщепление АДФ.
4. Ионсвязывающий центр фермента перемещается относительно толщины мембраны, в результате чего ион натрия оказывается на внешней стороне клетки.
5. Снаружи клетки вследствие уменьшения сродства фермента к ионам натрия и повышения сродства к калию происходит обмен этими ионами.
6. После отщепления фосфата фермент с присоединенными ионами калия снова изменяет положение относительно мембраны.
7. Ионы калия и неорганический фосфат высвобождаются в цитоплазму, и фермент возвращается в исходное состояние.
Таким образом, энергии, выделяющейся при гидролизе одной молекулы АТФ достаточно, чтобы вынести из клетки три иона натрия и внести два иона калия. Na+,K+ - насос способствует не только повышению градиентов концентраций ионов, но и возрастанию градиента электрического потенциала, то есть является электрогенным, так как сумма вносимых зарядов неравна сумме выносимых. Межклеточная среда приобретает «более положительный» заряд по сравнению с клеткой за счет выноса одного «лишнего» положительного иона.
В мембранах саркоплазматического ретикулума мышечных клеток и цитоплазматических мембранах кардиомиоцитов существует Ca2+ - насос, работа которого во многом сходна с механизмом переноса ионов Na+,K+ - насосом. За один цикл, в процессе которого расходуется одна молекула АТФ, переносится два иона кальция.
Активный транспорт протонов может осуществляться как с помощью подвижных переносчиков, так и через мембранные каналы. Протонные каналы представляют собой интегральные белки, образующие внутреннюю пору, где содержатся участки, к которым могут присоединяться протоны. Энергия АТФ расходуется на изменение конформации белковых молекул, вследствие чего сродство одних участков связывания к протонам понижается, а других – увеличивается, что заставляет протон перескочить на другой участок канала, сродство которого к протону на данный момент выше. Путем таких перескоков с одного участка связывания на другой ион водорода пересекает мембрану.
Перенос Н+ против градиентов их концентраций осуществляется не только за счет энергии, выделяющейся при гидролизе АТФ, но и за счет энергии фотонов. Этот способ используется галофильными бактериями, которые на свету выкачивают протоны из клетки, а энергию создавшегося градиента концентраций используют для синтеза АТФ.
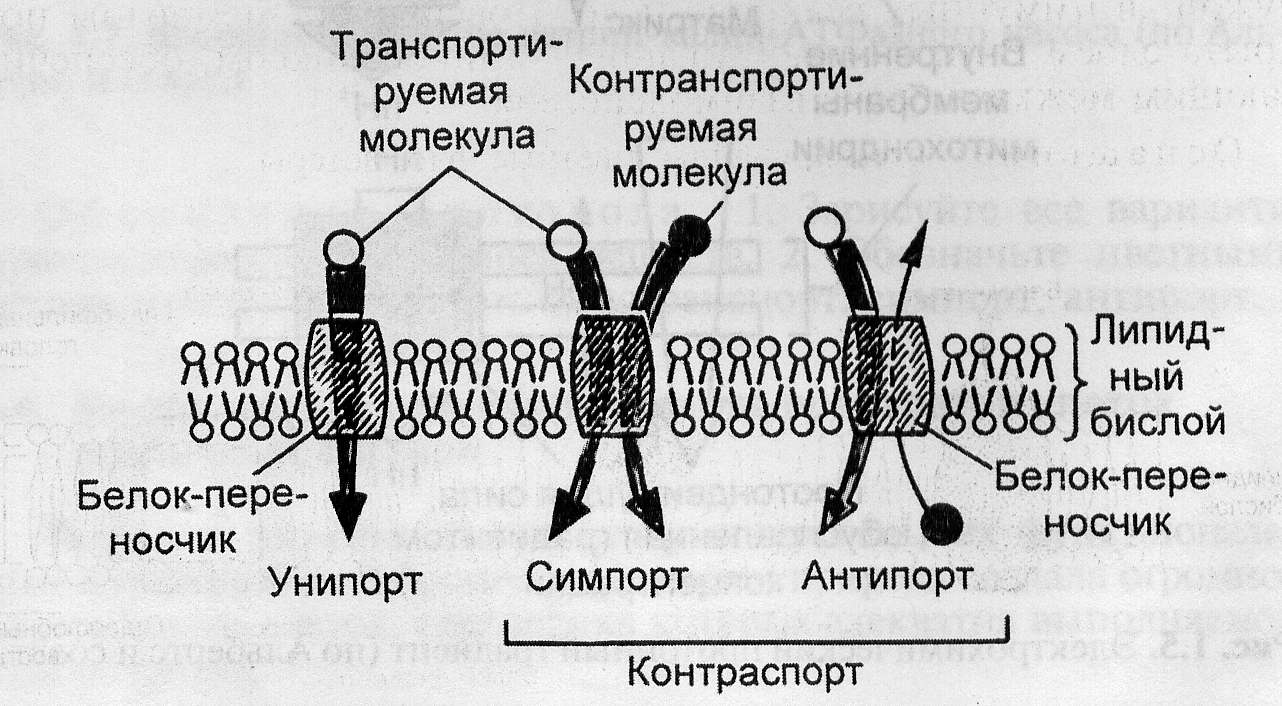
Вторично-активный транспорт. Транспорт, источником энергии которого служит не непосредственно АТФ или энергия окислительно-восстановительных реакций, а градиент концентраций других веществ, называется вторично-активным, или сопряженным транспортом. Существует три вида вторично-активного транспорта ионов: унипорт, симпорт и антипорт.
1. В случае унипорта за счет существования на мембране градиента электрического потенциала осуществляется однонаправленный транспорт заряженных частиц в сторону меньшего значения потенциала. Например, митохондрии в процессе своего функционирования активно выкачивают протоны из матрикса в цитоплазму, в результате чего их внутренняя область оказывается заряженной более отрицательно, чем внешняя. В нормальных условиях созданный градиент электрохимического потенциала используется для синтеза АТФ.
2. В процессе антипорта осуществляется транспорт одинаково заряженных ионов двух типов в разные стороны.
3. По механизму симпорта осуществляется транспорт противоположно заряженных ионов в одну сторону. При этом транспорт одного из ионов осуществляется по градиенту концентраций, а транспорт второго - по градиенту электрического потенциала, создаваемого транспортом первого иона.
Рис. 3. Схема вторично-активного транспорта веществ.
Одним из наиболее изученных случаев вторично-активного транспорта незаряженных молекул является всасывание глюкозы в стенках кишечника. Концентрация глюкозы в энтероцитах выше, чем в просвете кишечника, поэтому пассивное её всасывание невозможно. Клетки кишечника активно выкачивают натрий из энтероцитов в серозную область кишечника, в результате чего концентрация Na+ в клетке снижается по сравнению с его концентрацией снаружи. Благодаря этому становится возможным пассивный транспорт натрия из просвета кишечника в энтероцит. Однако простая диффузия ионов через бислой мало вероятна и натрий может пересечь мембрану только с помощью переносчика. При этом переносчик связывается не только с натрием, но и с молекулой глюкозы. Образовавшийся комплекс пассивно по градиенту концентраций ионов натрия и электрического потенциала переносится внутрь клетки. Таким образом, транспорт глюкозы осуществляется против градиента концентрации, но не за счет энергии АТФ, а за счет существования градиента концентрации другого вещества, в данном случае – ионов натрия. Подобные системы транспорта существуют и для многих других веществ, например, углеводов и аминокислот, что очень важно, так как для них отсутствуют специфические насосы.
| <== предыдущая лекция | | | следующая лекция ==> |
| Меры и правила безопасности при работе с ядовитыми | | | Понятие об электрическом поле. Силовая и энергетическая характеристики электрического поля |
Дата добавления: 2016-04-02; просмотров: 8137;