Биофизика фотосинтеза
Фотосинтез– это весь комплекс процессов ассимиляции фотонов и химических субстратов в растительном организме, которые приводят к его росту и развитию. Кроме этого, в фотосинтез входят: процессы синтеза пигментов, обеспечивающих поглощение фотонов и первичные фотохимические реакции; синтез соединений, отвечающих за формирование органов растительного организма и его приспособление к условиям окружающей среды. В более узком смысле фотосинтез – это синтез углеводов под действием видимого света. Термин фотосинтез был предложен Чарльзом Рейдом Бансом из Чикагского университета в начале прошлого столетия. В Европе этот процесс часто называют ассимиляцией или ассимиляцией углерода.
Многие живые организмы способны к фотосинтезу. Даже в кожном слое млекопитающих идет под действием света синтез химических соединений. Но основными объектами исследования фотосинтеза и практического использования его продуктов являются растения, водоросли и микроорганизмы. У высших растений основная масса синтезируемых соединений производится в листьях. В других частях также идут фотосинтетические процессы, управляющие развитием цветков, семян, изменением формы растений и их пространственной ориентацией.
Структурная основа для фотосинтетических процессов. Фотосинтетические процессы в листьях локализованы в специальных органеллах, называемых хлоропластами. Внутреннее пространство хлоропласта заполнено пластинчатыми гранулами зеленого цвета – гранами. Граны представляют собой стопки сильно сплюснутых мешочков – тилакоидами. Стенки тилакоидов – ламеллы – представляют собой тонкие фосфолипидные мембраны, внутри которых находятся липотропные участники первичных процессов фотосинтеза. К ним относятся белки, пигменты, переносчики электронов и протонов. Внутреннее межмембранное пространство тилакоидов является водной средой, в которой содержатся водорастворимые участники процесса. В межгранальном – стромальном – пространстве (также водном), находятся менее упакованные отдельные тилакоиды и ламеллы.
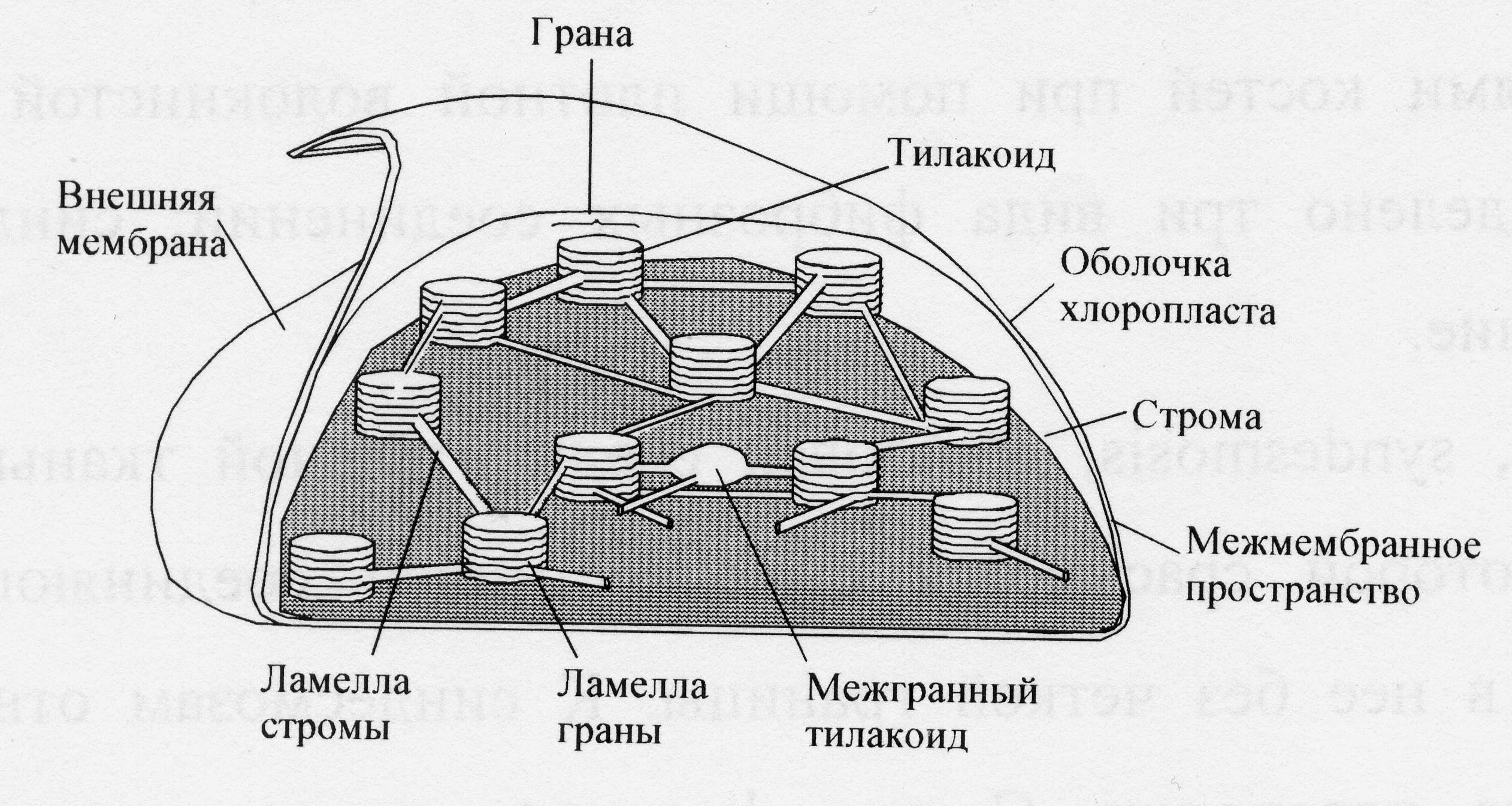
Рис.1. Строение хлоропласта.
У фотосинтезирующих бактерий нет хлоропластов, а участники процесса фотосинтеза расположены в мембранах, которые подобны ламеллам высших растений. Эти мембраны расположены под внешней оболочкой микроорганизма и образуют замкнутое пространство. Замкнутая топология мембранных образований и в хлоропластах, и в бактериях – важное условие, поскольку при функционировании фотосинтетического аппарата на этих мембранах, как и в митохондриях, возникает электрохимический потенциал для протонов.
Физико-химическая основа фотосинтеза. Основная продукция фотосинтеза - это соединения с высоким восстановительным потенциалом – углеводы. Рабочим телом фотосинтетической машины в высших растениях и некоторых водорослях является вода. Процесс фотосинтеза включает в себя две стадии: 1. Фотолиз, физическая сущность которого заключается в разложении воды с помощью квантов света на кислород и водород, причем кислород является в этом процессе побочным продуктом; и 2. Восстановление – получения глюкозы. Основным уравнением фотосинтеза в высших растениях является
6CO2 + 6H2O + Nhν = 6O2 + C6H12O6 + ∆G,
то есть, реакция идет только за счет энергии света и в итоге на один шаг реакции в её продуктах запасается ∆G= 686 ккал свободной энергии. Однако, во-первых, это уравнение объединяет два этапа превращения субстратов: световой и темновой, и, во-вторых, в данном уравнении не видны важные промежуточные продукты данной реакции. Поэтому для понимания необходимо записать эти этапы раздельно:
1. 6CO2+12H2O*+48hν+12НАДФ+18АДФ+18Ф → 6CO2+ 12НАДФН2+ 18АТФ + 6 O*2 →
2. → C6H12O6 + 6 O*2 + 6H2O + 12НАДФ +18АДФ +18Ф.
Теперь видно, что продуктом световой стадии является восстановленные переносчики НАДФН2 и АТФ, которые в темновой стадии восстановления углекислоты до глюкозы (цикл Кальвина) соответственно окисляются и гидролизуются. Звездочкой отмечен кислород, который получается при фоторазложении воды. Необходимо отметить, что вода, которая образуется в темновой стадии, содержит другой кислород.
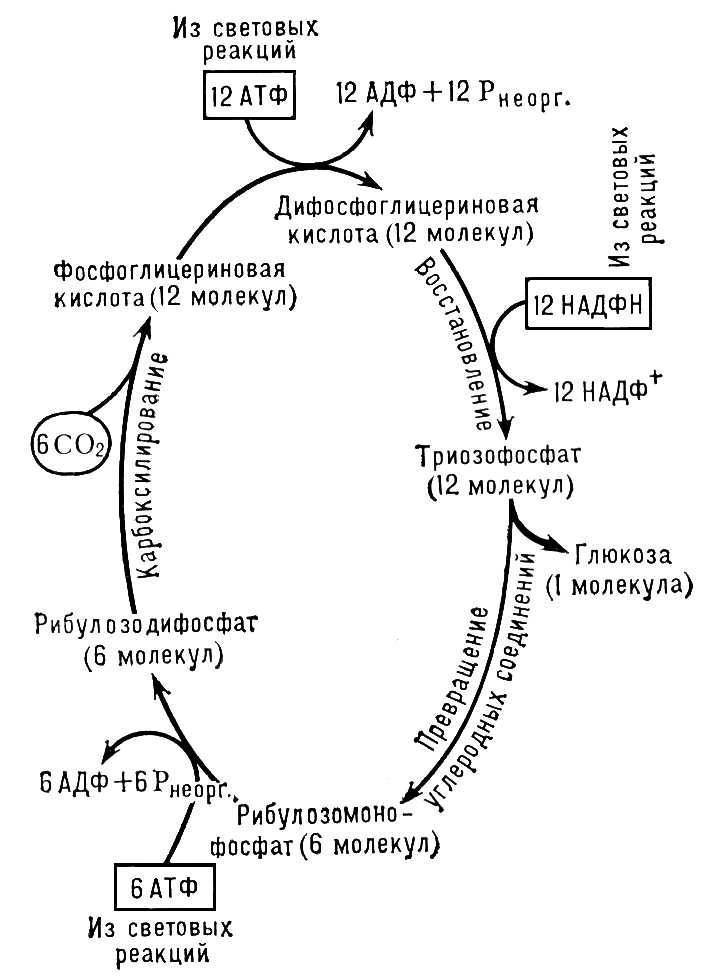
Рис.2. Схема фотосинтетических биохимических реакций.
Первая стадия фотосинтеза протекает на свету с помощью фотокатализатора. Фотокатализатором, или точнее, сенсибилизатором, в растениях является хлорофилл. Хлорофилл имеет гидрофобную часть и гидрофильную. То есть хлорофилл обладает амфотерными свойствами, которые способствуют самосборке агрегатов хлорофилла на границе липид-вода и на поверхности белков, где чередуются гидрофильные и гидрофобные участки. Структура центральной части хлорофилла аналогична гему железа в молекуле гемоглобина или цитохромах и обеспечивает поглощение света в видимом диапазоне. Существует три вида хлорофилла: a,b и c. Спектры поглощения хлорофиллов качественно похожи: в красной области – в районе 660-680 нм, в синей области – в районе длин волн 450 нм. Существование нескольких химически и физически различающихся форм хлорофиллов необходимо для того, чтобы сгладить спектр действия фотосинтеза в видимом участке солнечного спектра.
Красная граница фотосинтеза определяется самыми длинноволновыми формами хлорофилла, и у высших растений располагается в районе 700 нм. Свет в зеленой части спектра, поглощают некоторые пигменты, в частности, каротин, так как хлорофиллы имеют относительно малое поглощение в этой части спектра. Кроме этого каротин защищает хлорофилл от деструктивных процессов, которые возникают при длительном нахождении электрона на триплетном уровне.
В хлоропластах высших растений имеются две фотосистемы – фотосистема 1 (ФС1) и фотосистема 2 (ФС2), которые различаются по составу белков и пигментов. Светособирающая антенна ФС1 поглощает свет с длиной волны 700-730 нм, а ФС2 – 680 – 700 нм. Индуцированное светом окисление реакционных центров двух фотосистем сопровождается их обесцвечиванием, которое характеризуется изменениями спектров поглощения. Две фотосистемы связаны посредством цепи электронных переносчиков.
ФС2 является источником электронов для ФС1. Индуцируемое светом разделение зарядов в фотореакционных центрах обеспечивает перенос электрона от воды, разлагаемой в ФС2, к конечному акцептору электрона – молекуле НАДФ+. Цепь электронного транспорта, соединяющая две фотосистемы, в качестве переносчиков электрона включает в себя молекулы пластохинона, отдельный электрон-транспортный белковый комплекс (так называемый b/f-комплекс) и водорастворимый белок пластоцианин (Рс).
В ФС2 от возбужденного центра Р+ 680 электрон переносится сначала на первичный акцептор феофетин (Рhе), а затем на молекулу пластохинона Qа, прочно связанную с одним из белков ФС2. Затем электрон переносится на вторую молекулу пластохинона Qb, а Р+ 680 получает электрон от первичного донора – молекулы хлорофилла. После двукратного срабатывания реакционного центра ФС2 молекула пластохинона Qb получает два электрона и становится отрицательно заряженной. Эта молекула обладает высоким сродством к протонам, которые она захватывает из стромального пространства. После протонирования восстановленного пластохинона образуется электрически нейтральная форма этой молекулы (QbН2 или просто QН2), которая называется пластохинолом. Пластохинол выполняет роль подвижного переносчика двух электронов и двух протонов: покинув ФС2, молекула QН2 может легко перемещаться внутри тилакоидной мембраны, обеспечивая связь ФС2 с другими электрон-транспортными комплексами. Окислительный реакционный центр ФС2 обладает исключительно высоким сродством к электрону, то есть является очень сильным окислителем. Благодаря этому в ФС2 происходит разложение воды. Входящий в состав ФС2 водорасщепляющий комплекс (ВРК) содержит в своем активном центре группу ионов марганца, которые служат донорами электронов для Р+ 680 . Отдавая электроны окисленному реакционному центру Р+ 680 , ионы марганца становятся «накопителями» положительных зарядов, непосредственно участвующих в реакции окисления воды. В результате последовательного четырехкратного срабатывания реакционного центра ФС2 в Mn-содержащем активном центре ВРК накапливаются четыре сильных окислительных эквивалента (четыре «дырки») в форме окислительных ионов марганца, которые, взаимодействуя с двумя молекулами воды, катализируют реакцию её разложения. Таким образом, в результате последовательной передачи электронов от ВРК к Р+ 680 происходит синхронное разложение сразу двух молекул воды, сопровождающееся выделением одной молекулы кислорода и четырех протонов, которые диффундируют во внутритилакоидное пространство хлоропластов.
Образовавшаяся при функционировании ФС2 молекула пластихинола QН2 диффундирует внутри липидного бислоя тилакоидной мембраны к b/f-комплексу. При столкновении она связывается с ним и передает ему два электрона, при этом два освободившихся протона выходят во внутри тилакоидное пространство. В свою очередь b/f -комплекс служит донором электрона для пластоцианина, который выполняет роль связующего звена между b/f-комплексом и ФС1. Пластоцианин – это водорастворимый белок, имеющий в составе своего активного центра ион меди. Реакция восстановления и окисления пластоцианина сопровождается изменением валентности иона меди. Молекула пластоцианина быстро перемещается внутри тилакоида, обеспечивая перенос электрона от b/f-комплекса к ФС1. От восстановленного пластоцианина электрон поступает непосредственно к окислительным центрам ФС1. Таким образом, в результате совместного действия ФС1 и ФС2 два электрона от молекулы воды, разлагаемой в ФС2, через цепь электронного транспорта переносятся на молекулу НАДФ+, обеспечивая образование сильного восстановителя НАДФ•Н. Схематически выше описанные процессы представлены на рисунке 1.
Рис.3. Расположение электрон-транспортных комплексов (ФС1, ФС2, b/f-комплекса) и их взаимодействие в тилакоидной мембране.
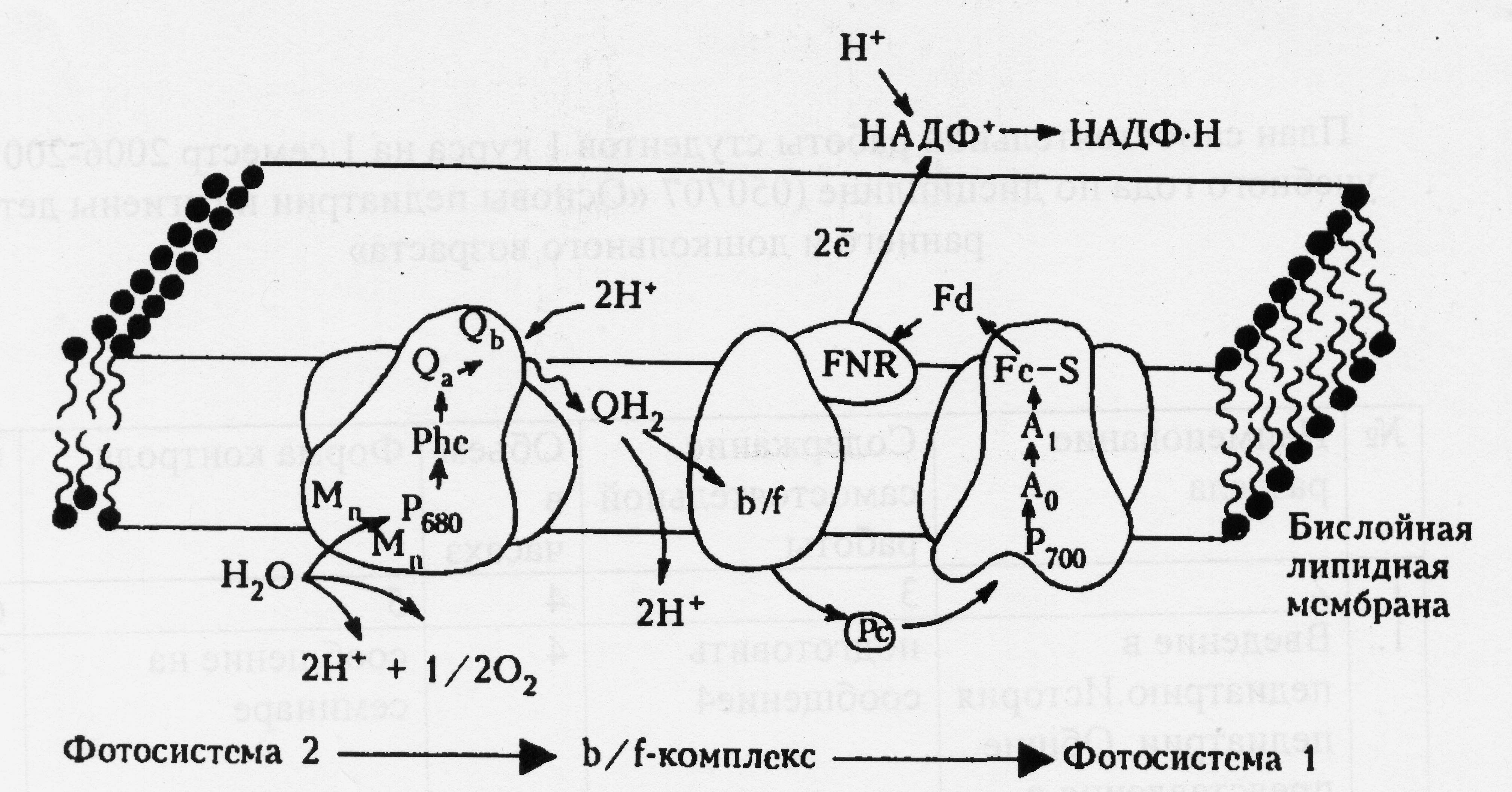
В состав ФС1 входят: Pc- пластоцианин, P700 – энергетическая ловушка и реакционный центр, первичный акцептор электрона – молекула хлорофилла (А0), вторичные акцепторы – молекула филлохинона (А1) и три переносчика белковой природы (ферредоксин), у некоторых в активном центре находятся атомы железа и серы.
В состав ФС2 входят: фотореакционный центр Р680, первичный акцептор – феофетин (Phe), вторичные акцепторы – молекулы пластохинона (Qa и Qb) и водорасщепляющий комплекс. В переносе электрона от акцепторов ФС1 к НАДФ+ участвуют растворимый в строме белок ферредоксин (Fd) и связанный с мембраной специальный электрон-транспортный комплекс ферредоксин - НАДФ+ - редуктаза (FNR), функционирующие на внешней стороне тилакоидной мембраны. При восстановлении одной молекулы НАДФ+ до НАДФ•Н на неё переносятся два электрона и один ион водорода, который захватывается из стромы.
Зачем хлоропластам нужны две системы? Известно, что фотосинтезирующие бактерии, которые используют в качестве донора электрона для восстановления окисленных реакционных центров различные органические и неорганические соединения (например, Н2S), успешно функционируют с одной фотосистемой. Дело в том, что энергии одного кванта видимого света недостаточно для эффективного прохождения электроном всего пути по цепи молекул-переносчиков от воды к НАДФ+.
Преобразование энергии в фотосинтетических системах. В настоящее время считается, что ФС1 ведет своё происхождение от зеленых бактерий, а ФС2 – от пурпурных. После того как в ходе эволюции ФС2 «включилась» в единую цепь электронного транспорта (ЦЭТ) вместе с ФС1, стало возможным преодолеть большую разницу окислительно-восстановительных потенциалов таких пар, как «кислород – вода» и «НАДФ+ - НАДФ•Н».
Перенос электронов по ЦЭТ сопровождается снижением энергии. Этот процесс можно уподобить самопроизвольному движению тела по наклонной плоскости. Уменьшение уровня энергии электрона в ходе его движения вдоль ЦЭТ не означает, что перенос электрона является энергетически бесполезным процессом. В нормальных условиях функционирования хлоропластов большая часть энергии, выделяющаяся в ходе электронного транспорта, не пропадает бесполезно, а используется для работы специального энергообразующего комплекса клетки, называемого АТФ-синтетазой. Этот комплекс катализирует энергетически невыгодный процесс образования АТФ из АДФ и неорганического фосфата. Поэтому принято говорить, что энергодонорные процессы электронного транспорта сопряжены с энергоакцепторными процессами синтеза АТФ.
Важную роль в обеспечении энергетического сопряжения в мембранах тилакоидов играют процессы протонного транспорта. Синтез АТФ тесно связан с переносом через АТФ-синтетазу трех протонов из тилакоидов в строму. Этот процесс становится возможным потому, что из-за асимметричного расположения переносчиков в мембране функционирование ЦЭТ хлоропластов приводит к накоплению избыточного количества протонов внутри тилакоида: ионы водорода поглощаются снаружи на стадиях восстановления НАДФ+ и образования пластохинола и выделяются внутри тилакоидов на стадиях разложения воды и окисления пластохинола. Следовательно, продуктами световой стадии фотосинтеза является энергия химических соединений АТФ и НАДФ, которая представляет собой запас энергии света. Эти продукты световой стадии фотосинтеза используются в темновых стадиях для образования органических соединений из углекислого газа и воды. Основными этапами преобразования энергии включают в себя следующие процессы:
1. Поглощение энергии света пигментами светособирающей антенны;
2. Перенос энергии возбуждения к фотореакционному центру;
3. Окисление фотореакционного центра и стабилизация разделенных зарядов;
4. Перенос электрона по цепи электронного транспорта, образование НАДФ•Н;
5. Трансмембранный перенос протонов;
6. Синтез АТФ.
Основные свойства фотосинтетического аппарата растений:
1. Насыщение световой кривой фотосинтеза с ростом освещенности:
Рис.4. По вертикальной оси – скорость ассимиляции углекислоты V, по горизонтальной оси – освещенность листа L.
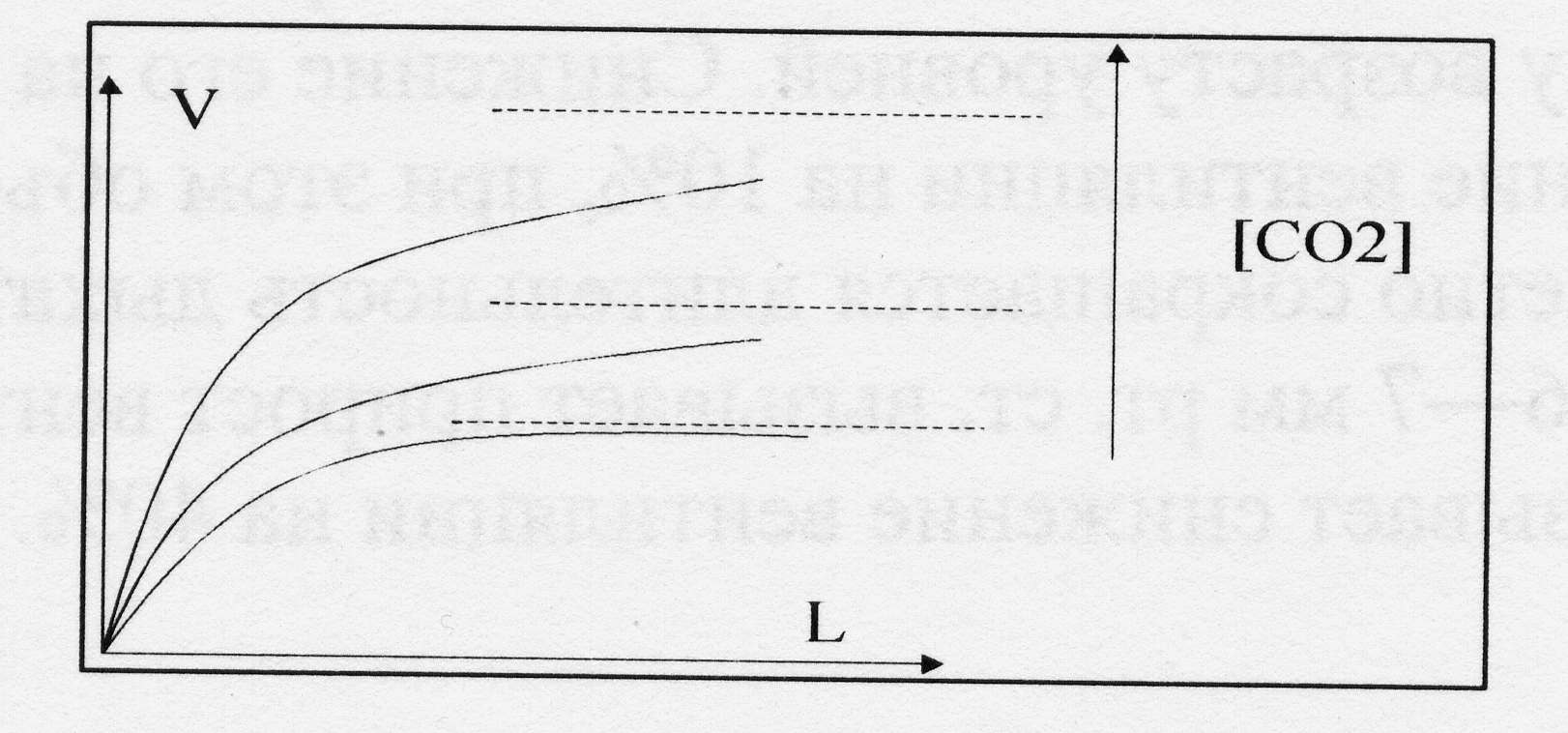
На графике видно, что кривые роста скорости фотосинтеза имеют выраженное насыщение. Предельное значение насыщения растет с ростом концентрации CO2 в воздухе. Причиной этого явления являются ограничения скорости потока электронов в электрон-транспортной цепи, когда скорость их отвода на CO2 лимитируется его диффузией из окружающей среды.
2. «Эффект усиления» Эммерсона. Если измерять интенсивность фотосинтеза от двух монохроматических источников света с длинами волн λ1 < 690 нм и λ2 > 690 нм, то она окажется в несколько раз больше, чем интенсивность фотосинтеза при свете той же мощности, но содержащем лишь одну длину волны λ1 или λ2. Это объясняется тем, что для нормального протекания фотосинтеза необходимо сбалансированное возбуждение фотосистем ФС1 и ФС2. В противном случае поток электронов ослабевает. «Память» «эффекта усиления» составляет 1 секунду. Если освещать лист растения поочередно светом с λ1 и λ2 с интервалом меньше 1 секунды, то скорость фотосинтеза будет неотличима от скорости при одновременном облучении светом с λ1 и λ2 . Это время характеризует время удержания пластохиноном избытка или дефицита электронов между ФС1 и ФС2.
3. Изменение окислительно-восстановительного состояния цитохромов b и f при освещении светом с разными длинами волн. При освещении фотосинтетического аппарата светом с λ1 < 690 нм цитохромы восстанавливаются, а при освещении светом с λ2 > 690 нм цитохромы окисляются. Это происходит потому, что свет с длиной волны λ1 возбуждает преимущественно ФС2, направляющую электроны к цитохромам, а свет с длиной волны λ2 возбуждает в основном ФС1, отсасывающую электроны от цитохромов.
4. Эффект Кока-Жолио. Если зеленый лист растения выдержать в темноте около часа, а затем начать освещать короткими вспышками света, то выход кислорода будет иметь следующий вид

Рис.5. Количество кислорода, выделяемого на одну вспышку. N – номер вспышки. На две первые вспышки кислород практически не выделяется, на третью вспышку выделение максимально, на четвертую несколько меньше, далее серия повторяется с периодом 4, и через 20-30 вспышек модуляция сглаживается, и выделения становятся одинаковыми. Эффект объясняется дискретностью работы кислород-выделяющего комплекса. Если вспышка достаточно короткая, так что кислород-выделяющий комплекс успевает получить от Р680 за вспышку только одну дырку, то последовательное одноэлектронное окисление реакционного центра ФС2 изменяет каждый раз суммарную валентность ионов Mn на единицу. А так как не все реакционные центры получают возбуждения во время каждой вспышки, то со временем состояния кислород-выделяющих комплексов разсинхронизируются и суммарный эффект сглаживается.
5. Задержанная флуоресценция. Вслед за затуханием флуоресценции молекулы хлорофилла (несколько наносекунд) в живом листе наблюдается слабое остаточное свечение с характерным временем затухания от нескольких секунд до минуты.
6. Индукция флуоресценции. При включении освещения наблюдается флуоресценция с определенным значением квантового выхода. Через некоторое время начинается рост уровня флуоресценции до нового значения квантового выхода.
Одноквантовые схемы первичных стадий в фотосинтезе некоторых бактерий.
· У серных бактерий всего одна фотосистема: электроны забираются из H2S и восстанавливают НАДФН с последующим образованием сахаров, то есть цикл переноса электронов не замкнут. Такие бактерии не выделяют кислород.
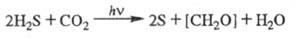
· У некоторых несерных пурпурных бактерий фотосистема осуществляет только замкнутый цикл переноса электронов.
· Солелюбивые бактерии запасают энергию непосредственно в виде электрохимического потенциала протонов и последующего синтеза АТФ с помощью, так называемого, бактериородопсинового фотосинтеза.
| <== предыдущая лекция | | | следующая лекция ==> |
| Ремни безопасности неработоспособны или имеют видимые надрывы на лямке. | | | НОРМА И ДЕВИАЦИЯ В СЕКСУАЛЬНОМ ПОВЕДЕНИИ В КОНТЕКСТЕ РАЗЛИЧНЫХ ПОДХОДОВ |
Дата добавления: 2016-04-02; просмотров: 6033;