Пищеварение в тонкой кишке
В тонкой кишке осуществляется перемешивание кислого химуса со щелочными секретами поджелудочной железы, кишечных желез и печени, деполимеризация питательных веществ до конечных продуктов (монометров), способных поступать в кровоток, продвижение химуса в аборальном направлении, экскреция метаболитов и др.
Пищеварение в этом отделе начинается в полости кишки {по
лостное пищеварение), а затем продолжается в зоне гликокаликса и
исчерченной каймы энтероцитов слизистой кишечника {пристеночное
пищеварение). Как полостное, так и пристеночное пищеварение осу
ществляется ферментами секретов поджелудочной железы и
кишечного сока под влиянием желчи.
Секреторная деятельность поджелудочной железы.Поджелудочная железа — большая железа смешанной секреции. Эндокринный отдел поджелудочной железы, представленный клетками островков Лангерганса, продуцирует ряд гормонов (инсулин, глюкагон и др.) непосредственно в кровь. Экзокринный отдел представлен панкре-оцитами оцинусов, секретирующими пищеварительные ферменты, а также рацинозными эпителиоцитами и клетками вставочных отделов поджелудочной железы мелких выводных протоков, секретирующих воду, карбонаты, электролиты. Образующийся панкреатический сок поступает через систему выводных протоков в двенадцатиперстную кишку. Состав и свойства панкреатического сока зависят от количества и качества пиши.
Состав и свойства поджелудочного сока. В сутки у человека вырабатывается 1,5-2,5 л панкреатического сока, изотоничного плазме крови, щелочной реакции (рН 7,5-8,8). Такая реакция обусловлена содержанием ионов бикарбоната, которые обеспечивают нейтрализацию кислого желудочного содержимого и создают в двенадцатиперстной кишке щелочную среду, оптимальную для действия панкреатических ферментов. Кроме того, в состав панкреатического сока входят катионы Na+, K+, Са2+ М2+ и анионы
412
Cl- , HCO32- , HPO42- , а также слизистые вещества. Концентрация бикарбонатов в соке прямо пропорционально зависит от скорости его секреции. Между концентрацией бикарбонатов и хлоридов зависимость обратно пропорциональная.
Поджелудочный сок содержит ферменты для гидролиза всех видов питательных веществ: белков, жиров и углеводов.
Белки расщепляют протеолитические ферменты, которые по механизму гидролиза разделяются на две группы: эндопептидазы (трипсин, химотрипсин, эластаза) расщепляют внутренние пептидные связи белков, образуя пептиды и аминокислоты; экзопептидазы (карбоксииеп-тидаза А и В и аминопептидаза) расщепляют в белках и пептидах конечные связи, освобождая аминокислоты одну за другой. Панкре-атоциты выделяют протеолитические ферменты в виде неактивных проферментов трипсиногенов, химотрипсиногенов, прокарбоксипепти-даз А и В, которые активируются в двенадцатиперстной кишке.
Трипсиноген превращается в активный трипсин при отщеплении гексапептида ферментом энтерокиназой (продукта энтероцитов Брун-неровых желез). После образования трипсина процесс активации его предшественника при рН 6,8-8,0 становится аутокаталитическим. Образовавшийся активный трипсин, в свою очередь, становится активатором для химотрипсиногена, прокарбоксипептидаз А и В, проэластазы.
В соке поджелудочной железы содержатся липолитические ферменты, которые выделяются в неактивном (профосфолипаза А) и активном состоянии (панкреатическая липаза, лецитиназа).
Панкреатическая липаза гидролизует нейтральные жиры до жирных кислот и моноглицеридов, фосфолипаза А расщепляет фосфо-липиды до жирных кислот. Гидролиз жиров липазой усиливается в присутствии желчных кислот и ионов кальция.
Амилолитический фермент сока (панкреатическая альфа- амилаза) расщепляет крахмал и гликоген до ди- и моносахаридов. Дисаха-риды далее под влиянием мальтазы и лактазы превращаются в моносахариды.
Нуклеотические ферменты принадлежат к фосфодиэстеразам. В панкреатическом соке они представлены рибонуклеазой (гликолиз рибонуклеиновой кислоты) и дезоксинуклеазой (гидролиз дезокси-нуклеиновой кислоты).
С целью предотвращения самопереваривания те же клетки, которые секретируют протеолитические ферменты, одновременно секре-тируют вещество, называемое ингибитором трипсина. Это вещество накапливается в цитоплазме железистых клеток, окружая гранулы фермента, что предупреждает активацию трипсина как внутри секреторных клеток, так и в ацинусах и протоках поджелудочной железы. Поскольку трипсин активирует остальные панкреатические протеолитические ферменты, его ингибитор также предупреждает их последующую активацию.
Калликреин, активируемый трипсином, биологически активное вещество, стимулирует образование в крови каллидина, гипотензивного пептида, идентичного брадикинину.
413
Секреция ионов бикарбоната и воды. В отличие от ферментов ионы бикарбоната и воды секретируются в больших количествах эпителиальными клетками протоков поджелудочной железы. Обильная секреция панкреатического сока сопровождается значительным увеличением концентрации ионов бикарбоната — до 145 Мэкв/л, что в пять раз превышает их содержание в плазме. Такое многократное увеличение бикарбоната в секрете обеспечивается тесно взаимосвязанными процессами, начинающимися с того, что двуокись углерода диффундирует внутрь клеток из крови и под влиянием карбоангидразы взаимодействует с водой, образуя угольную кислоту, диссоциирующую на ионы бикарбоната и водорода. Бикарбонат проходит через мембрану клетки в просвет протоков. Ионы водорода, образовавшиеся при диссоциации угольной кислоты внутри клетки, активно обмениваются на ионы натрия, которые транспортируются в панкреатический проток. Движение ионов натрия и бикарбоната из крови в просвет протока создает осмотический градиент, вызывающий движение воды в панкреатический проток и образование раствора бикарбоната.
Регуляция панкреатической секреции.Секреторные клетки поджелудочной железы вне периода пищеварения находятся в состоянии покоя и отделяют сок лишь в связи с периодической деятельностью желудочно-кишечного тракта. Секреция поджелудочной железы происходит иод воздействием нервных влияний и гуморальных раздражителей, возникающих при поступлении пищи в пищеварительный тракт, атакже при виде, запахе пищи и в случае действия привычной обстановки ее приема. Как и в случае желудочной секреции, процесс отделения поджелудочного сока разделяется на три фазы: сложнорефлекторную (мозговую или цефалическую), желудочную и кишечную.
Сложнорефлекторная фаза секреции начинается не только при непосредственном воздействии элементов пиши на рецепторы ротовой полости, но и при виде, запахе пищи и при действии той обстановки, в которой происходит акт еды. Поступление пищи в полость рта и глотки вызывает рефлекторное возбуждение, наслаивающееся на уже начавшуюся секрецию поджелудочной железы.. Дуга рефлекса включает рецепторы полости рта, чувствительные нервные волокна, идущие в продолговатый мозг, центральные парасимпатические нейроны, эфферентные волокна вагуса, секреторные клетки поджелудочной железы. Эта фаза значительно менее выражена, нежели в предыдущих отделах пищеварительного тракта.
Желудочная фаза секреции является следствием поступления пищи в желудок. Возбуждение поджелудочной железы возникает при механическом, химическом и гуморальном раздражении рецепторов желудка. Афферентные импульсы, возникающие в результате раздражения хеморецепторов слизистой оболочки желудка, по чувствительным нервным волокнам поступают в центральную нервную систему, откуда по блуждающему нерву эфферентные влияния направляются к поджелудочной железе. Химическими раздражителями являются как
414
вещества, вырабатывающиеся в самом желудочно-кишечном тракте, так и содержащиеся в пище. Натуральными раздражителями, вызывающими возбуждение поджелудочной железы, являются НС1, овощные соки, жиры и продукты их гидролиза. Гуморальным регулятором поджелудочной железы в этой фазе является гормон антраль-ного отдела желудка гастрин. Последний, всасываясь в кровь, возбуждает секрецию поджелудочной железы.
Кишечная фаза секреции начинается после поступления химуса в двенадцатиперстную кишку. В это время вырабатывается большая часть панкреатического сока. Количество и состав секрета поджелудочной железы зависит от качества и количества пищи, контролируется рецептивными клетками кишечника и, в первую очередь, двенадцатиперстной кишки. Эта фаза секреции развивается под влиянием рефлекторных влияний и кишечных гормонов. Доказано существование мощных дуоденопанкреатических рефлексов. Общность иннервации поджелудочной железы, двенадцатиперстной кишки и печени с желчными ходами обусловливает их функциональную взаимосвязь.
Поступление хлористоводородной кислоты и продуктов переваривания пищи в верхний отдел тонкой кишки стимулирует секрецию поджелудочной железы. Стимуляция продолжается при поступлении желчи в двенадцатиперстную кишку и новых частиц переваренной пищи в тонкую кишку. Однако поджелудочную железу в этой фазе секреции преимущественно стимулируют кишечные гормоны секретин и холецистокинин. Секретин высвобождается при действии НС1 на S-клетки двенадцатиперстной кишки. Под его влиянием вырабатывается большое количество сока поджелудочной железы, богатого бикарбонатами и бедного ферментами, так как он преимущественно действует на функции эпителиальных клеток протоков и почти не влияет на панкреатоциты. Холецистокинин действует преимущественно на ацинарные клетки, вызывая секрецию панкреатического сока, богатого ферментами. Высвобождение холецистокинина из I-клеток слизистой двенадцатиперстной и тощей кишки в наибольшей мере стимулирует продукты начального гидролиза пищевого белка и жира, а также аминокислоты, в меньшей степени этот процесс стимулируется хлористоводородной кислотой и углеводами. Богатый ферментами панкреатический сок секретируется лишь при совместном действии на железу секретина и холецистокинина, по-тенциированных ацетилхолином.
Панкреатическую секрецию стимулируют также вазоактивный ин-тестинальный полипептид (ВИП), серотонин, образующийся в энте-рохромафинных клетках слизистой желудочно-кишечного тракта и ткани поджелудочной железы, инсулин, бомбезин, субстанция Р, соли желчных кислот. Тормозное влияние на секрецию оказывают глю-кагон, кальцитонин, ПП, соматостатин, ГИП. Эффекты интести-нальных пептидов опосредуются путем их влияния на секреторную активность желудочных желез, что сопровождается увеличением кислотности желудочного химуса, который при поступлении в двенадцатиперстную кишку стимулирует выделение ее гормонов.
415
Парасимпатические нервные влияния оказывают пусковое, а симпатические — тормозное влияние на железу. Нервные влияния здесь менее выражены, чем гуморальные. Из слизистой оболочки двенадцатиперстной кишки выделен полипептид, стимулирующий секрецию хи мотрипсиногена.
Снижение панкреатической секреции происходит при болевых раздражениях, во время сна, при напряженной физической и умственной работе.
В ответ на потребление белковой и углеводной пищи (мясо, хлеб) наблюдается резкое увеличение секреции в первые два часа, с максимумом отделения сока на втором часе после приема пищи. В этом случае продолжительность секреции может быть от 4- 5 часов (мясо) до 9-10 часов (хлеб). При приеме жирной пищи (молоко) максимальный подъем секреции имеет место на третьем часе, продолжительность секреции на этот стимул равна 5 часам (рис.9.4).
Роль печени в пищеварении.Печень является железой, в которой происходят многочисленные и сложнейшие биохимические процессы, обеспечивающие гомеостазис жизненно важных и тесно связанных с обменом веществ систем в организме. Она оказывает влияние на обмен белков, пептидов, углеводов, пигментный обмен, выполняет дезинтоксикационную (обезвреживающую) и желчеобразовательную функции.
Желчеобразовательная и желчевыделительная функции печени. Желчь является секретом и, одновременно, экскретом, постоянно вырабатываемым печеночными клетками-ге-патоцитами. Образование желчи происходит в печени посредством активного и пассивного транспорта воды, глюкозы, креатинина, электролитов, витаминов и гормонов через клетки и межклеточные пространства, а также активного транспорта желчных кислот клетками и реабсорбции воды, минеральных и органических веществ из желчных капилляров, протоков и желчного пузыря, в которых она наполняется продуктом муцинсекретирующих клеток.
Поступив в просвет двенадцатиперстной кишки, желчь включается в процесс пищеварения и участвует в смене желудочного пищеварения на кишечное, инактивируя пепсин и нейтрализуя кислоту содержимого желудка, создавая благоприятные условия для активности ферментов поджелудочной железы, особенно липаз. Желчные кислоты желчи эмульгируют жиры, снижая поверхностное натяжение капель жира, что создает условия для образования тонкодисперсных частиц, способных всасываться без предварительного гидролиза, способствуют увеличению его контакта с липолитическими ферментами. Желчь обеспечивает всасывание в тонкой кишке нерастворимых в воде высших жирных кислот, холестерина, жирорастворимых витаминов (Д, Е, К) и солей кальция, усиливает гидролиз белков и углеводов, а также всасывание продуктов их гидролиза, способствует ресинтезу триглицеридов в энтероцитах. Благодаря щелочной реакции желчь участвует в регуляции работы пилорического сфинктера. Она оказывает стимулирующее влияние на моторную деятельность
416
тонкой кишки, в том числе и на деятельность кишечных ворсинок, в результате чего повышается скорость абсорбции веществ в кишке; участвует в пристеночном пищеварении, создавая благоприятные условия для фиксации ферментов на кишечной поверхности. Желчь является одним из стимуляторов секреции поджелудочной железы, желудочной слизи, моторной и секреторной деятельности тонкой кишки, пролиферации и слущивания эпителиоцитов, а главное —желчеобразовательной функции печени. Наличие пищеварительных ферментов позволяет желчи участвовать в процессах кишечного пищеварения, она также предупреждает развитие гнилостных процессов, оказывая бактериостатическое действие на кишечную флору.
Секрет гепатоцитов представляет собой золотистую жидкость, почти изотоничную плазме крови, ее рН равен 7,8-8,6. Суточная секреция желчи у человека составляет 0,5-1,0 л. Желчь содержит 97,5% воды и 2,5% сухого остатка. Составными ее частями являются желчные кислоты, желчные пигменты, холестерин, неорганические соли (натрия, калия, кальция, магния, фосфаты, железо и следы меди). В желчи содержатся жирные кислоты и нейтральные жиры, лецитин, мыла, мочевина, мочевая кислота, витамины А,В,С, некоторые ферменты (амилаза, фосфатаза, протеаза, каталаза, оксидаза), аминокислоты, гликопротеиды. Качественное своеобразие желчи определяют ее основные компоненты: желчные кислоты, желчные пигменты и холестерин. Желчные кислоты — специфические продукты обмена веществ в печени, билирубин и холестерин имеют внепеченочное происхождение.
В гепатоцитах из холестерина образуются холевая и хенодезокси-холевая кислоты (первичные желчные кислоты). Соединяясь в печени с аминокислотами глицином или таурином, обе эти кислоты выделяются в виде натриевой соли таурохолевой кислоты. В дис-талъном отделе тонкой кишки около 20% первичных желчных кислот превращаются под действием бактериальной флоры во вторичные желчные кислоты — дезоксихолевую и литохолевую. Здесь же примерно 90-85% желчных кислот активно реабсорбируются, возвращаются по портальным сосудам к печени и включаются в состав желчи. Остальные 10- 15% желчных кислот, связанных, в основном, с непереваренной пищей, выводятся из организма, а их убыль восполняется гепатоцитами.
Желчные пигменты — билирубин и биливердин — являются экс-кретируемыми продуктами метаболизма гемоглобина и придают желчи ее характерную окраску. В желчи человека и плотоядных животных преобладает билирубин, который обусловливает ее золотисто-желтый цвет, а в желчи травоядных содержится биливердин, окрашивающий желчь в зеленый цвет. В гепатоцитах билирубин образует водорастворимые коньюгаты с глюкуроновой кислотой и, в незначительном количестве, с сульфатами. Из пигментов желчи образуются пигменты мочи и калауробилин, урохром и стеркобилин.
Секрет выделяется гепатоцитами в просвет желчных капилляров, из которых через внутридольковые или междольковые желчные ходы желчь поступает в более крупные желчные протоки, сопровожда-
417
ющие разветвления портальной вены. Желчные протоки постепенно сливаются и в области ворот печени образуют печеночный проток, из которого желчь может поступить либо через пузырный проток в желчный пузырь, либо в общий желчный проток.
Жидкая и прозрачная, золотисто-желтого цвета печеночная желчь при движении по протокам начинает претерпевать некоторые изменения в связи с всасыванием воды и добавлением муцина желчных путей, однако это существенно не изменяет ее физико-химических свойств. Наиболее значительные изменения в желчи происходят во внепищеварительный период, когда она направляется через пузырный проток в желчный пузырь. Здесь желчь концентрируется, становится темной, пузырный муцин способствует увеличению ее вязкости, нарастает удельный вес, всасывание бикарбонатов и образование солей желчных кислот приводит к снижению активной реакции (рН 6,0-7,0). В желчном пузыре за 24 часа желчь концентрируется в 7-10 раз. Благодаря такой концентрационной способности желчный пузырь человека, обладающий объемом лишь 50- 80 мл, может вмещать желчь, образующуюся в течение 12 часов (таблица 9.2).
Таблица 9.2 Состав печеночной и пузырной желчи
| Составные вещества и ионы | Печеночная желчь | Пузырная желчь |
| Вода | 97.5 Г/дл | 92 г/дл |
| Желчные соли | 1.1 г/дл | 6 г/дл |
| Билирубин | 0.04 г/дл | 0.3 г/дл |
| Холестерол | 0.1 г/дл | 0.3-0.9 г/дл |
| Жирные кислоты | 0.12 г/дл | 0.3-1.2 г/дл |
| Летицин | 0.04 г/дл | 0.3 г/дл |
| Na+ | 145 ммоль/л | 130 ммоль/л |
| К+ | 5 ммоль/л | 12 ммоль/л |
| Са++ | 5 ммоль/л | 23 ммоль/л |
| а- | 100 ммоль/л | 25 ммоль/л |
| нсо3- | 28 ммоль/л | 10 ммоль/л |
Во время периодической и пищеварительной деятельности желудочно-кишечного тракта желчь изливается через общий желчный проток в двенадцатиперстную кишку, где принимает участие в пищеварении.
Регуляция секреции и выделение желчи. Секреция желчи происходит непрерывно, независимо от того, находится ли пища в пищеварительном тракте или нет. Акт еды рефлекторно усиливает отделение желчи через 3-12 мин. Мощными пищевыми возбудителями секреции желчи являются желтки, молоко, мясо, хлеб. Наибольшее количество желчи образуется при потреблении смешанной пищи.
Желчеобразование изменяется при раздражении интероцепторов желудочно-кишечного тракта. К числу его гуморальных стимулято-
418 . .
ров относится сама желчь (механизм саморегуляции), а также секретин, повышающий отделение воды и электролитов (бикарбонатов), желчных солей и желчных пигментов. Желчеобразование стимулируют также глюкагон, гастрин, холецистокинин.
Неравные пути, по которым к печени поступают стимулирующие или тормозящие импульсы, представлены холинергическими волокнами блуждающих и диафрагмальных нервов и адренергическими волокнами симпатических нервов и сплетений. Блуждающий нерв усиливает выработку желчи, симпатический — тормозит.
Выделение желчи в двенадцатиперстную кишку зависит от тонуса гладких мышц внепеченочных желчных путей, деятельности мышц сфинктера и стенки желчного пузыря, а также сфинктера, находящегося в месте слияния пузырного и общего желчного протока и сфинктера, расположенного в месте впадения общего желчного протока в двенадцатиперстную кишку (сфинктера Одди).
Направленное движение желчи из печени в двенадцатиперстную кишку возникает вследствие разности давлений в начальном отделе системы желчевыделения, в желчных ходах, протоках и двенадцатиперстной кишке. Давление в желчных капиллярах является результатом секреторной активности гепатоцитов, а в ходах и протоках оно создается сокращениями гладкомышечной стенки, согласованными с моторной активностью сфинктеров протоков и желчног* пузыря и с перистальтической деятельностью двенадцатиперстнсй кишки.
Вне процесса пищеварения сфинктер общего желчного протока закрыт и желчь поступает в желчный пузырь. Во время пищеварения желчный пузырь сокращается, сфинктер общего желчного гро-тока расслабляется и желчь поступает в двенадцатиперстную юшку. Такая согласованная деятельность обеспечивается рефлекторными и гуморальными механизмами. При поступлении пищи в пищеварительный тракт возбуждается рецепторный аппарат ротовой полости, желудка, двенадцатиперстной кишки. Сигналы по афферентным нервным волокнам поступают в центральную нервную систему и оттуда по блуждающему нерву к мышцам желчного пузыря и сфинктера Одди, вызывая сокращение мышц пузыря и расслабление сфинктера, что обеспечивает выделение желчи в двенадцатиперстную кишку.
Основным гуморальным стимулятором сократительной активности желчного пузыря является холецистокинин. Он вызывагт одновременное сокращение пузыря и расслабление сфинктера Одди, в результате чего желчь поступает в двенадцатиперстную кишку.
В клинической практике при исследовании сократительной функции желчного пузыря используются в качестве стимуляторов желчевыделения жидкое масло, яичный желток, пилокарпин, питуитрин, ацетилхолин, гистамин, сульфат магния.
Метаболические функции печени. Выполняя в организме роль биохимической лаборатории, печень оказывает выраженное влияние на обмен белков, жиров и углеводов. Благодаря синтезу альбуминов (12-15 г в сутки) она обеспечивает нормальное
419
онкотическое давление во внутренней среде организма и транспорт кровью плохо растворимых в воде веществ. Печенью же синтезируется почти 90% богатых липопротеидами а1- глобулинов, также участвующих в транспорте веществ, и 75% а2- глобулинов. В этой фракции представлено значительное количество глико- и липопротеидов, содержатся церулоплазмин, а2- антитромбин, гаптоглобин, а2- макроглобулин. Печенью образуется более 50% fi- глобулинов, во фракции которых имеются гемопексин, трансферрин, B2-микроглобулин, значительное количество липопротеидов. Она оказывает влияние на механизм свертывания крови, обеспечивая его фибриногеном.
Значительна роль печени и в преобразовании жиров, в частности, жирных кислот с короткой цепью в высшие жирные кислоты. Из синтезируемых ею липопротеидов холестерин служит источником синтеза желчных кислот и гормонов.
Постоянной функцией печени является синтез и расходование гликогена, что обеспечивает поддержание углеводного гомеостаза — стабильную гликемию. Снижение уровня глюкозы в плазме крови, которое может сопровождать возросшие энергетические потребности организма, приводит к выделению гормонов надпочечников и поджелудочной железы — адреналина и глюкагона, что сопровождается гликогенезом в печени и выведением глюкозы в кровь. Часть глю-кюзы используется самой печенью для синтеза жирных и желчных кислот, стероидных гормонов и гликопротеидов.
Способность печени предохранять организм от действия токсических веществ — дезинтоксикационная, барьерная функция — имеет существенное значение для сохранения жизни организма посредством биотрансформации веществ эндогенного и экзогенного происхождения. Эти вещества могут быть результатом гниения белка в толстом кишечнике (индол, фенол, скатол), образовываться в процессе обмена веществ или поступать в организм с пищей извне (ксенобиотики). Реакции биотрансформации могут осуществляться посредством гидроксилирования (перенос атома кислорода на молекулу "чужеродного" вещества с образованием гидроксильной группы), конъюгации (соединение "чужеродного" вещества с молекулой глю-куроновой, серной кислоты или же с метильной группой; пример — индолсернистые, скатолсернистые, фенолсернистые нетоксичные парные эфиры), специфического действия ферментных систем. Из ферментов наибольшее значение имеют глютатионтрансфераза, которая осуществляет биотрансформацию путем преобразования "чужеродного" вещества с помощью глутатиона, исполняет роль белка-лиган-дина, транспортирующего "чужеродное" вещество в отделы гепато-цита, где оно преобразуется.
Мочевинообразование — одна из важнейших дезинтоксикационных функций печени, заключающаяся в преобразовании ядовитых осколков белковой молекулы в нетоксичное вещество. При дезаминиро-вании аминокислот, нуклеотидов и других азотистых соединений печень принимает участие в синтезе аммиака, превышение предельно допустимых концентраций которого становится высокотоксичным для организма. Дезинтоксикация аммиака происходит посредством
420
использования его для синтеза мочевины. Даже при удалении 90-95% ткани печени сохраняется функция дезаминирования и синтеза мочевины.
Очень тесно с функцией печени связан метаболизм гормонов. Она осуществляет инактивацию адреналина, норадреналина, дофамина, альдостерона, серотонина, гастрина, эстрогенов и андрогенов. Особенно велика ее роль в метаболизме эстрогенов.
Секрецияжелез тонкой кишки.Кишечный сок является продуктом деятельности бруннеровых, либеркюновых желез и клеток всей слизистой оболочки тонкой кишки, представляет собой мутную, вязкую жидкость. В сутки у человека выделяется до 2,5 л кишечного сока. Выделение ферментов кишечного сока в принципе отличается от секреции ферментов другими пищеварительными железами. Секретирующие клетки слюнных, желудочных и поджелудочной желез выделяют пищеварительный сок и сохраняют свою целостность, а отделение кишечного сока связано с гибелью железистых клеток. Б слизистой оболочке тонкой кишки происходит, с одной стороны, интенсивное новообразование клеток, а с другой — непрерывное слущивание, отторжение погибших клеток с образованием слизистых комочков, поэтому при центрифугировании кишечный сок разделяется на жидкую и плотную части.
Жидкая часть кишечного сока образуется водными растворами органических и неорганических веществ, главным образом поступающими из крови, и небольшого количества содержимого разрушенных клеток кишечного эпителия. Неорганические вещества, содержащиеся в жидкой части сока, в основном — хлориды, бикарбонаты и фосфаты натрия, калия, кальция; органические вещества — белки, аминокислоты, мочевина и другие продукты обмена веществ организма. рН секрета составляет 7,2-7,5, при интенсивной секреции рН сока повышается до 8,6.
Плотная часть кишечного сока имеет вид слизистых комков желтовато-серого цвета, состоит из разрушенных эпителиальных клеток, их энзимов и слизи (секрет бокаловидных клеток) и обладает большей ферментной активностью, чем жидкая.
Ферменты кишечного сока. В кишечном соке более 20 ферментов, принимающих участие в пищеварении. Они гидроли-зуют пептиды и пептоны белков, расщепленные в желудке, до аминокислот, жиры —до глицерина и жирных кислот, а углеводы — до моносахаридов. В кишечном соке присутствуют пептидазы: амино-полипептидазы, дипептидазы, лейцинаминпептидазы и др., объединенные общим названием — эрипсины. Протеолитический фермент секрета бокаловидных клеток кишечника человека — ингибин. Расщепление нуклеотидов и нуклеиновых кислот в кишечном соке осуществляется нуклеотидазой и нуклеазой.
Липолитическими ферментами тонкокишечного сока являются липаза, фосфолипаза, холестеролэстераза. Амилолитические ферменты кишечного сока: амилаза, лактаза, сахараза. Особое место занимает гамма-амилаза, которая обладает специфическими особеннос-
421
тями, она прочно связана с липопротеиновой мембраной эпители-оцитов и практически не десорбируется в полость кишечника. Гамма-аминаза участвует в расщеплении полисахаридов и завершает гидролиз декстринов и олигосахаридов, которые образуются при гидролизе крахмала. Важным ферментом в кишечном соке является энтерокиназа, активирующая панкреатический трипсиноген.
Пищеварение в тонкой кишке является трехзвеньевой системой ассимиляции пищи: полостное пищеварение — мембранное пищеварение — всасывание.
Полостное пищеварение в тонкой кишке осуществляется за счет пищеварительных секретов и их энзимов, которые поступают в полость тонкой кишки (панкреатический секрет, желчь, кишечный сок) и действуют на пищевое вещество, прошедшее ферментную обработку в желудке. Крупномолекулярные вещества гидролизируют-ся до олигомеров по типу полостного пищеварения. Под влиянием ферментов гидролиз олигомеров завершается в зоне гликокаликса и микроворсинок эпителиоцитов.
Ферменты, осуществляющие мембранное пищеварение, имеют различное происхождение. Часть их адсорбируется из полости тонкой кишки, куда они поступают в составе поджелудочного и кишечного соков. Эти ферменты связаны с гликокаликсом микроворсинок. Другие ферменты переносятся из энтероцитов и фиксируются на цитоплазматических мембранах микроворсинок. Те энзимы, которые адсорбируются на мембранах микроворсинок энтероцитов, имеют более длительное время полезной работы, чем те, которые работают в полости кишечника.
Регуляция секреции кишечного сока. Секреторные клетки желез слизистой оболочки тонкой кишки возбуждаются при местном воздействии: секреция осуществляется в месте нахождения пищевого комка, как результат влияния механических раздражителей, которые вызывают выделение большого количества слизи, а сок при этом содержит малое количество ферментов. Мощным химическим раздражителем секреторных клеток желез слизистой оболочки тонкой кишки являются продукты переваривания белка желудочным соком, панкреатический сок, жирные кислоты, молочный сахар.
Специфическое действие химических раздражителей получило название ферментного приспособления. Действие каждого химического раздражителя вызывает выделение кишечного сока с определенным набором ферментов. Так, например, жирные кислоты стимулируют образование кишечными железами липазы, диета со сниженным содержанием белка приводит к резкому снижению активности эн-терокиназы в кишечном соке. Однако не все кишечные ферменты участвуют в процессах специфического ферментного приспособления. Выработка пептидаз не претерпевает существенных изменений даже при резком недостатке белка в течение 5 месяцев. Образование липазы в слизистой оболочке кишечника не меняется ни при повышенном, ни при пониженном содержании жира в пище. Таким образом, имеются как адаптирующиеся ферменты, так и ферменты, не участвующие или слабо участвующие в процессах адаптации.
422
В основе ответной реакции желез слизистой оболочки кишечника на раздражение механо-и хеморецепторов лежит рефлекторный механизм. Возбуждение, возникающее в рецепторах слизистой оболочки тонкой кишки, передается по чувствительным волокнам в центральную нервную систему, откуда поступают регулирующие стимулы к железистому аппарату кишечника. Парасимпатические влияния усиливают кишечную секрецию, симпатические — тормозят.
Деятельность желез кишечника стимулируется гормонами энтери-новой системы: ГИП, ВИП, мотилином; соматостатин тормозит их активность. Энтерокринин и дуокринин, выделяющиеся из слизистой оболочки кишечника, стимулируют либеркюновы и бруннеровы железы.
На деятельность желез слизистой оболочки кишечника оказывают также влияние гормоны общего действия, в частности, гормоны коры надпочечников, которые активируют образование адаптируемых кишечных ферментов, способствуют более полной реализации специфических нервных импульсов, регулирующих интенсивность выработки различных ферментов. Кортикостероиды вызывают увеличение секреции энтерокиназы, при этом выделение альфа-глюкозидазы и пептидаз не меняется.
Двигательная функция тонкой кишки и ее регуляция. Кислый химус из желудка поступает в тонкий кишечник, где под влиянием сократительной деятельности гладкомы-шечной оболочки тонкой кишки пищевые вещества продолжают измельчаться, подвергаются перемешиванию с щелочными пищеварительными соками и желчью, продвигаются вдоль кишки. Благодаря двигательной активности осуществляется смена пристеночного слоя химуса и повышается внутриклеточное давление. Таким образом, моторная деятельность кишечника обеспечивает эффективный гидролиз пищи, всасывание пищеварительных веществ и передвижение содержимого по пищеварительной трубке в аборальном направлении.
Моторика кишечника осуществляется посредством функциональных единиц гладкомышечной ткани (пучками мышечных клеток диаметром около 100 мкм), формируется благодаря координированным сокращениям и расслаблениям наружного продольного и внутреннего циркулярного слоя мышц. В кишечнике имеются датчики ритма (пейсмекерные зоны), задающие частоту активности соседним участкам. Одна зона находится в области сфинктера Одди в двенадцатиперстной кишке, вторая — в подвздошной кишке.
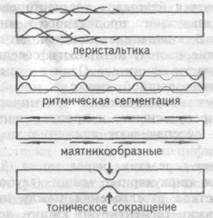
Четыре основных типа кишечных сокращений — ритмическая сегментация, перистальтические (непропульсивные и пропульсивные), маятникообразные и тонические — обеспечивают движение химуса по кишке.
Ритмическая сегментация возникает вследствие чередования (8- 10 раз в минуту) участков сокращения циркулярных мышц (глубокие перетяжки 1-2 см ширины) и участков расслабления между ними (15-20 см ширины) (рис.9.5). Перетяжки сегментируют кишку, делят ее содержимое на части, способствуют повышению давления в полости сегмента, приводящее к растяжению его в участке расслабле-
423
 ния. Как только в зоне расслабления возникает участок наибольшего растяжения, миоциты его возбуждаются, что сопровождается образованием новой перетяжки. В это время мышцы ранее образованной перетяжки расслабляются и участок растягивается. Новый сегмент формируется из частей двух смежных, а его содержимое является смесью частей их объемов.
ния. Как только в зоне расслабления возникает участок наибольшего растяжения, миоциты его возбуждаются, что сопровождается образованием новой перетяжки. В это время мышцы ранее образованной перетяжки расслабляются и участок растягивается. Новый сегмент формируется из частей двух смежных, а его содержимое является смесью частей их объемов.
Перистальтические сокращения подобны волне, распространяющейся в аборальном направлении и продвигающей содержимое вдоль по кишке. Эта волна возникает в результате образования перехвата при сокращении циркулярных мышц выше комка химуса и расширения полости кишки при сокращении продольной мускулатуры ниже комка. Непропульсивная перистальтика — волна сокращения, распространяющаяся на короткие расстояния, ей не предшествует расслабление. Пропульсивная перистальтика — волны сокращения различной силы, распространяющиеся со скоростью от 0,1-3 см/с до 7-21 см/с, на разные расстояния. Этим волнам обычно предшествуют волны расслабления. Слабые волны приводят к перемещению только поверхностных слоев химуса. Мощные, стремительные волны могут распространяться вдоль всего тонкого кишечника вплоть до илеоцекальной заслонки. Несколько таких одновременных сокращений почти полностью освобождают тонкий кишечник от пищевых масс. Такой тип перистальтических сокращений обычно наблюдается к концу пищеварения в кишечнике.
Значительная часть волн начинается в двенадцатиперстной кишке или в момент перехода содержимого из желудка в кишку, или на несколько секунд раньше, чем волна сокращения желудка достигает пилорического сфинктера. Вдоль кишки могут одновременно осуществляться несколько перистальтических сокращений.
Маятникообразные движения возникают при доминирующем сокращении продольного слоя с участием циркулярного. Такие движе-

 Характер моторики Место Функция
Характер моторики Место Функция

|
| желудок тонкая кишка | транспорт и перемешивание | |
| тонкая кишка толстая кишка | перемешивание | |
| тонкая кишка толстая кишка | контакт химуса со слизистой оболочкой | |
| сфинктеры | функциональное разделение отделов |
 Рис.9.5. Виды движений кишечника.
Рис.9.5. Виды движений кишечника.
ния приводят, главным образом, к смешению кишечной стенки относительно химуса и перемещению его "вперед-назад" со слабым поступательным движением в аборальном направлении.
Частота маятникообразных сокращений и ритмической сегментации в одном и том же участке кишки приблизительно одинакова. Чередование ритмической сегментации и маятникообразных движений способствует тщательному перемешиванию пищевых масс.
Тонические сокращения обусловлены определенным исходным (ба-залъным) тонусом гладких мышц тонкой кишки, могут передвигаться с малой скоростью и вызывать одновременное повышение тонуса кишки, суживая ее на большом протяжении. Они являются основой перистальтического, маятникообразного сокращения и ритмической сегментации. Отсутствие тонуса мышц кишечника (атония) при парезах исключает возможность любого вида сокращений. Тонические сокращения характерны для кольцевых сплетений гладкомышечных волокон- сфинктеров.
Функционально значимыми являются также сокращение и расслабление ворсинок кишечника, которые происходят постоянно, в течение всего процесса пищеварения. Благодаря ритмичным сокращениям ворсинок они соприкасаются с новыми порциями химуса, что способствует всасыванию и оттоку лимфы по центральным лимфатическим сосудам.
Моторная активность тонкой кишки зависит от механических свойств и химического состава ее содержимого. Если пища содержит грубые частицы (изделия из муки грубого помола, отруби, овощи, фрукты с высоким содержанием клетчатки) или жиры, то двигательная активность кишки возрастает. На моторику кишечника оказывают также стимулирующее действие слабые растворы кислот, щелочи и соли.
Регуляция моторики тонкой кишки обеспечивается механизмами миогенного происхождения, нервными интрамуральными влияниями и центральной нервной системой, гормонами энтерального и неэн-терального происхождения.
Миогенные механизмы регуляции. В основе моторной деятельности кишок лежат свойства гладкомышечных клеток спонтанно сокращаться и отвечать сокращением на раздражение, в том числе на растяжение. Физиологическое свойство гладких мышц —способность к автоматии — обусловлено спонтанной периодической деполяризацией мембраны клеток в виде циклического процесса. Циклические ионные сдвиги и изменения уровня мембранного потенциала приводят к ритмическому появлению медленных электрических волн, возникновению на них плато потенциалов действия и к мышечному сокращению.
К миогенным механизмам регуляции моторики тонких кишок относится также сократительная реакция клеток их гладких мышц на растяжение. Сокращение продольных мышц обеспечивает растяжение циркулярных до уровня, достаточного для их сокращения. Продольный слой гладких мышц является пусковым механизмом (датчиком ритма) сокращений для циркулярного слоя мышц. Рас-
425
 пространение возбуждения из продольного на циркуляторный слой происходит по соединяющим эти слои мышечным пучкам.
пространение возбуждения из продольного на циркуляторный слой происходит по соединяющим эти слои мышечным пучкам.
Деятельность датчиков ритма и других миогенных механизмов регуляции моторики кишки (уровень мембранного потенциала, величина разрядов пиковых потенциалов и др.), а также скорость проведения по ней медленных волн, зависят от уровня обмена веществ, содержания биологически активных веществ и температуры. Снижение температуры тела или участка, где локализован датчик ритма, приводит к снижению частоты сокращений медленных волн и скорости их проведения.
Нервная регуляция двигательной деятельности тонкого кишечника осуществляется интрамуральной и центральной нервной системой (вегетативный отдел спинного мозга и ствола, гипоталамус, лимби-ческая нервная система и в определенной мере — кора больших полушарий головного мозга) (рис.9.6.).
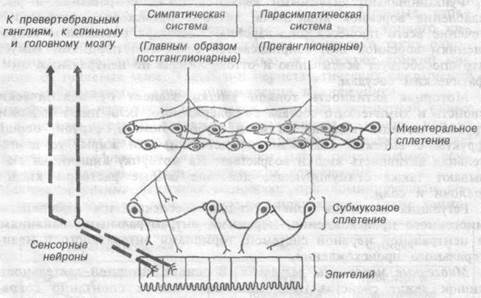
Рис.9.6. Центральные и периферические механизмы нервной регуляции деятельности тонкой кишки.
 Экстрамуральные нервы (блуждающие и чревные) — эффекторное звено вегетативного отдела центральной нервной системы —выполняют функцию передачи тормозных и возбуждающих влияний на моторную деятельность кишечника, и кроме того, содержат нервные волокна для передачи сенсорной информации из кишки в нервные центры. Посредством этих нервов обеспечивается рефлекторная регуляция моторики кишечника. В составе блуждающих и чревных нервов содержатся возбуждающие и тормозные волокна. Симпатические волокна преимущественно тормозят, а парасимпатические — возбуждают со-
Экстрамуральные нервы (блуждающие и чревные) — эффекторное звено вегетативного отдела центральной нервной системы —выполняют функцию передачи тормозных и возбуждающих влияний на моторную деятельность кишечника, и кроме того, содержат нервные волокна для передачи сенсорной информации из кишки в нервные центры. Посредством этих нервов обеспечивается рефлекторная регуляция моторики кишечника. В составе блуждающих и чревных нервов содержатся возбуждающие и тормозные волокна. Симпатические волокна преимущественно тормозят, а парасимпатические — возбуждают со-
426
крашения тонкой кишки. Реакция кишки на раздражение блуждающего нерва во многом зависит от исходного ее состояния в момент нанесения раздражения. Стимуляция периферического конца перерезанного блуждающего нерва на фоне слабых сокращений кишки вызывает усиление ее моторики, а на фоне активных ее сокращений — торможение. Блуждающие и чревные нервы содержат афферентные волокна, по которым импульсы из кишечника достигают различных уровней центральной нервной системы.
Основными моторными рефлексами кишечника являются:
1) пищеводно-кишечный рефлекс, который возникает при повыше
нии давления в пищеводе до 2- 10 мм рт.ст. и вызывает раздраже
ние его рецепторов. В результате повышается тонус и усиливаются
сокращения верхнего отдела тонкой кишки в случае, если кишка
находилась в состоянии покоя или слабых сокращений. На фоне
сильных или максимальных ее сокращений выраженного эффекта не
отмечается. Возбуждающие влияния в этом случае передаются по
блуждающим, а тормозные — по чревным нервам;
2) желудочно-кишечный моторный рефлекс наблюдается при раз
дражении механорецепторов желудка во время наполнения его пи
щей, что приводит к появлению или усилению имеющихся сокра
щений тонкой кишки и проксимальных отделов толстой кишки.
Рефлексы, возникающие в верхних отделах (двенадцатиперстная,
тощая) тонких кишок при раздражении желудка называются гастро-
дуоденальными или гастроеюнальными рефлексами, а усиление со
кращений толстой кишки при раздражении желудка — желудочно-
толстокишечным рефлексом. Возбуждение к тонкому кишечнику и
проксимальным отделам толстой кишки при раздражении желудка
передается по стенке пищеварительной трубки с участием интраму-
ральных нервных сплетений и рефлекторно — посредством экстра-
муральных нервов, главным образом, блуждающих, с замыканием
рефлекторной дуги в центральной нервной системе. Рефлекторное
возбуждение сокращений толстой кишки при раздражении желудка
передается по тазовым и блуждающим нервам. Блуждающие нервы
играют роль афферентного пути этого рефлекса, а эфферентные
пути проходят в тазовых нервах;
3) кишечно-кишечный моторный рефлекс — осуществляется при
адекватном механическом и химическом раздражении тонких кишок
и проявляется в возбуждении или усилении сокращений нижележа
щих отделов кишечника. Нервное возбуждение распространяется в
этом случае в каудальном направлении с участием интра- и экстра
муральной нервной системы.
Регуляция моторной деятельности кишечника осуществляется не только возбуждающими рефлекторными влияниями, но и тормозными, что позволило выделить следующие тормозные рефлексы:
1) рецепторное торможение кишечника, которое возникает во время еды и проявляется в торможении или снижении тонуса верхних отделов тонкой кишки, за которыми следует возбуждение ее моторной деятельности (это явление получило название воспринимающего торможения кишки — рецепторная релаксация — и имеет значение
427
для приема первых порций пищи в кишку из желудка). Афферентный путь этого рефлекса начинается возбуждением рецепторов глотки и корня языка, а эфферентные волокна представлены волокнами чревного нерва;
2) кишечно-кишечный тормозной рефлекс — вызывается сильным
раздражением любой части желудочно-кишечного тракта и сопро
вождается торможением сокращений других частей, за исключением
илеоцекального сфинктера. Важнейшая роль в осуществлении этого
рефлекса принадлежит чревным нервам. Замыкание рефлекса про
исходит в спинном мозге ниже ТVI;
3) ректо-энтеральный рефлекс — является следствием раздраже
ния прямой кишки и сфинктера ее ампулы,проявляется в виде
торможения толстой и тонкой кишки. Передача тормозных влияний
с прямой кишки на верхние участки тонкой осуществляется волок
нами ваго-симпатических стволов через спинной мозг.
Существуют тормозные кишечные рефлексы, замыкающиеся в брюшных ганглиях. В этом случае торможение моторики участков тонкой кишки может проявляться при раздражении толстой или других отделов тонкой кишки после полной их децентрализации. Эти висцеро-висцеральные рефлексы кишечника замыкаются в ганглиях энтеральной нервной системы (в узлах по ходу брыжеечных нервов, брюшных ганглиях), а также в узлах симпатической цепочки.
Основной закон рефлекторной регуляции моторной деятельности желудочно-кишечного тракта, которому подчиняется и тонкая кишка, сводится к следующему. Адекватное раздражение любого участка желудочно-кишечного тракта вызывает возбуждение моторики в раздражаемом и нижележащих участках и усиление продвижения содержимого в каудальном направлении от места раздражения; при этом тормозится моторика и задерживается прогрессивное продвижение химуса в вышележащих участках и отделах желудочно-кишечного тракта.
Таким образом, моторная деятельность любого участка кишки или отдела желудочно-кишечного тракта является суммарным результатом возбуждающего действия пищи на этот участок, а также возбуждающих влияний на него проксимальных и тормозных влияний с дистальных — относительно данного участка — отделов желудоч-но- кишечного тракта.
Адреналин и норадреналин, действуя на альфа- и бета-адренер-гические рецепторы, в основном, тормозят моторную деятельность кишки. Большие дозы ацетилхолина вызывают двухфазную реакцию: возбуждение, сменяющееся торможением. В малых дозах ацетилхо-лин возбуждает сокращение кишки. От уровня адреналина и ацетилхолина в крови зависят моторные ответы кишок при рефлекторных влияниях на них. Повышение уровня адреналина в крови усиливает тормозные и ослабляет возбуждающие нервные влияния на моторику кишки. Ацетилхолин вызывает противоположный эффект.
В регуляции моторной деятельности желудочно-кишечного тракта играют роль высшие отделы центральной нервной системы — гипоталамус, структуры лимбической системы и кора больших полуша-
428
рий головного мозга. Стимуляция ядер передних и средних отделов гипоталамуса преимущественно возбуждает, а заднего — тормозит моторику тонкой и толстой кишки.
Гипоталамус активирует деятельность коры мозга, представляя собой "пейсмекерный" механизм всего пишевого поведения. Раздражение передней части поясной извилины (лимбическая область коры) вызывает усиление сокращений тела желудка, торможение сокращений его пилорического отдела и возбуждение сокращений тонкой кишки.
О корковом контроле деятельности пищеварительного тракта свидетельствуют результаты прямого электрического раздражения полей коры больших полушарий мозга. Страх, испуг, опасность, беспокойство, боль вызывают торможение моторики желудочно-кишечного тракта. Сильные эмоции и длительный страх сопровождаются бурной моторикой кишечника и диарреей ("нервный понос").
Дата добавления: 2016-03-27; просмотров: 895;