ОСМОТИЧЕСКОЕ РАЗВЕДЕНИЕ И КОНЦЕНТРИРОВАНИЕ МОЧИ
Способностью к образованию мочи с большей осмотической концентрацией, чем кровь, обладают лишь почки теплокровных животных. Многие исследователи пытались разгадать физиологический механизм этого процесса, но лишь в начале 50-х годов XX века была обоснована гипотеза, согласно которой образование осмотически концентрированной мочи связано с механизмом противоточно-поворотной множительной системы некоторых участков нефрона.
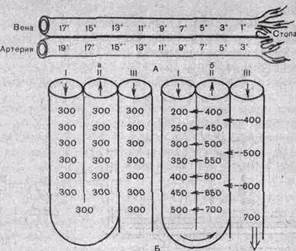
Принцип противоточного обмена достаточно широко распространен в природе и используется в технике. Механизм работы такой системы рассмотрим на примере кровеносных сосудов в конечностях арктических животных. Во избежание больших потерь тепла кровь в параллельно расположенных артериях и венах конечностей течет таким образом, что теплая артериальная кровь согревает охлажденную венозную кровь, движущуюся к сердцу (рис. 204). В стопу притекает артериальная кровь с низкой температурой, что резко уменьшает теплоотдачу. Здесь такая система функционирует только как противоточный обменник: в почке же она обладает множительным эффектом. Для лучшего понимания ее работы рассмотрим систему, состоящую из трех параллельно расположенных трубок. Трубки I и II дугообразно соединены на одном из концов (рис. 204, Б). Стенка, общая для обеих трубок, обладает способностью переносить соли, но она не пропускает воду. Когда в такую систему через вход 1 наливают жидкость с концентрацией 300 мосмоль/л и она не течет, то через некоторое время в результате транспорта солей в трубке I жидкость станет гипотонической, а в трубке 11 — гипертонической. В том случае, когда жидкость течет по трубкам непрерывно, начинается кон-центрирование солей. На каждом горизонтальном уровне перепад их концентраций вследствие одиночного эффекта транспорта солей не может превышать 200 мосмоль/л, однако по длине трубки происходит умножение одиночных эффектов и система начинает работать как противоточная множительная система. Так как по ходу движения жидкости из нее извлекаются не только соль, но и некоторое количество воды, концентрация раствора все более повышается по мере приближения к изгибу петли. В трубке III регу-
| Рис.204. Схема функционирования противоточно-поворотной системы. А — теплообменник в сосудистой системе конечностей арктических животных; обмен тепла между артериальной и венозной кровью способствует сбережению тепла и на каждом уровне не превышает 1° — 2° С. Б — модель противоточной множительной системы в начальном состоянии (а) и в период эффективного концентрирования мочи (б). Объяснение в тексте. |
|
| ^— Активный транспорт Nа <^—| Пассивный транспорт НдО ^й-а Пассивный транспорт мочевины |
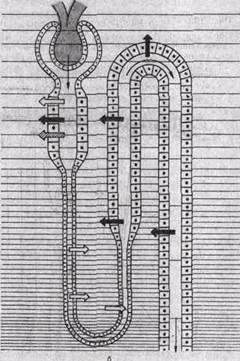
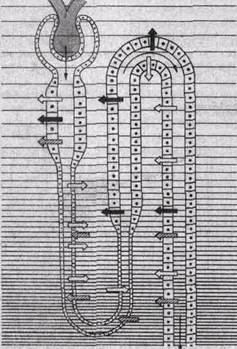
Рис.205. Повышение концентрации (показано штриховкой увеличенной частоты) осмотически активных веществ в различных участках почки.
а - состояние антидиуреза; б — состояние водного диуреза. Широкими стрелками обозначено направление транспорта основных веществ, участвующих в осмотическом концентрировании; тонкими стрелками — двн жение первичной и вторичной мочи.
лируется проницаемость стенок для воды; когда стенка начинает пропускать воду, объем жидкости в ней уменьшается. При этом вода идет в сторону большей осмотической концентрации. В результате этого растет концентрация жидкости в трубке III и уменьшается объем содержащейся в ней жидкости. Концентрация в ней веществ будет зависеть от ряда условий, ^ том числе от работы противоточной множительной системы трубок I и II. Как будет ясно из последующего изложения, работа почечных канальцев в процессе осмотического концентрирования мочи похожа на описанную модель.
В зависимости от состояния водного баланса организма почки выделяют разведенную или концентрированную мочу. В процессе осмотического концентрирования мочи в почке принимают участие все отделы канальцев, сосуды мозгового вещества, интер-стициальная ткань. Из 100 мл фильтрата, образовавшегося в клубочках, ^з его реаб-сорбируются к концу проксимального сегмента. Оставшаяся в канальцах жидкость содержит осмотически активные вещества в такой же концентрации, как и ультрафильтрат плазмы крови, хотя и отличается от него по составу вследствие реабсорбции ряда веществ в предшествующих частях нефрона. Далее канальцевая жидкость переходит из коркового слоя почки в мозговое вещество — в нисходящий (тонкий) отдел петли нефрона (петля Генле) и движется до вершины почечного сосочка, где каналец изгибается на 180°, и моча переходит в восходящий отдел петли, расположенный параллельно ее нисходящему отделу.
Функциональное значение различных отделов петли неоднозначно. Когда жидкость из проксимального отдела канальца поступает в тонкий нисходящий отдел петли нефрона, она попадает в зону почки, в интерстициальной ткани которой концентрация осмотически активных веществ выше, чем в коре почки. Это повышение осмолярной концентрации в наружной зоне мозгового вещества обусловлено деятельностью толстого восходящего отдела петли нефрона. Его стенка непроницаема для воды, а клетки транспортируют ионы С1~ и Nа+ в интерстициальную ткань. Стенка нисходящего отдела петли проницаема для воды, и поэтому вода всасывается из просвета канальца в окружающую межуточную ткань почки по осмотическому градиенту, а осмотически активные вещества остаются в просвете этого отдела канальца.
Чем дальше от коры по продольной оси находится жидкость в нисходящем колене петли, тем выше ее осмолярная концентрация. В каждых соседних участках нисходящего отдела петли имеется лишь небольшое нарастание осмотического давления, но по длине петли осмолярная концентрация постепенно растет от 300 мосмоль/л почти до 1450 мосмоль/л. Иначе говоря, на вершине петли нефрона осмолярная концентрация жидкости возрастает в несколько раз и при этом объем ее уменьшается. При дальнейшем передвижении жидкости по восходящему отделу петли нефрона происходит реабсор-бция ионов С1~ и Nа+, вода остается в просвете канальца, поэтому в начальные части дистального извитого канальца всегда поступает гипотоническая жидкость, концентрация осмотически активных веществ в которой менее 200 мосмоль/л.
Из гипотонической жидкости по осмотическому градиенту реабсорбируется вода, осмолярная концентрация жидкости в этом отделе увеличивается, т. е. жидкость в просвете канальца становится изоосмотической. Окончательное концентрирование мочи происходит в собирательных трубках; они расположены параллельно канальцам петли нефрона, в мозговом веществе почки. Как отмечалось выше, в интерстициальной жидкости мозгового вещества почки возрастает осмолярная концентрация. Вследствие этого из жидкости собирательных трубок реабсорбируется вода и концентрация мочи в них увеличивается, уравновешиваясь со все повышающейся осмолярной концентрацией внутреннего мозгового вещества почки. В конечном счете выделяется гиперосмотическая моча, в которой максимальная концентрация осмотически активных веществ может быть равна осмолярной концентрации интерстициальной жидкости на вершине почечного сосочка (рис. 205).
В условиях дефицита воды в организме усиливается секреция антидиуретического гормона гипофиза (АДГ), что увеличивает проницаемость стенок конечных частей дистального сегмента и собирательных трубок для воды.
14 -Физиология человека417
В отличие от наружной зоны мозгового вещества почки, где повышение осмолярности основано главным образом на транспорте хлоридов, увеличение осмолярной концентрации во внутренней зоне мозгового вещества почки зависит от нескольких механизмов. Особую роль в осмотическом концентрировании играет накопление мочевины. Стенки проксимального канальца проницаемы для мочевины. В этом отделе нефрона реабсорбируется до 50% профильтровавшейся мочевины. Однако при извлечении жидкости из извитого дистального канальца оказалось, что содержание мочевины даже несколько превышает ее количество, поступившее с фильтратом, и составляет около 1 !0%. Было показано, что имеется система внутрипочечного кругооборота мочевины, которая участвует в осмотическом концентрировании мочи. В просвете собирательных трубок вследствие реаб-сорбции воды повышается концентрация мочевины, АДГ увеличивает проницаемость собирательных трубок в мозговом веществе не только для воды, но и для мочевины. Когда увеличивается проницаемость канальцевой стенки для мочевины, она диффундирует в мозговое вещество почки. Постоянное поступление во внутреннее мозговое вещество мочевины, ионов С1~ и Nа+, реабсорбируемых клетками тонкого восходящего отдела петли нефрона и собирательных трубок, обеспечивает повышение осмотической концентрации в мозговом веществе почки. Вслед за увеличением осмолярности окружающей собирательные трубки межуточной ткани возрастает и реабсорбция воды из них и повышается эффективность осморегулирующей функции почки. Изменение проницаемости канальцевой стенки для мочевины позволяет понять, почему очищение от мочевины уменьшается при снижении мочеотделения.
Прямые кровеносные сосуды мозгового вещества почки, подобно канальцам петли нефрона, также образуют противоточную систему, играющую очень важную роль в осмотическом концентрировании. Благодаря особенностям расположения прямых сосудов обеспечивается эффективное кровоснабжение мозгового вещества почки, но не происходит вымывания осмотически активных веществ, поскольку в крови прямых сосудов наблюдаются такие же изменения осмотической концентрации, как и в тонком нисходящем отделе петли нефрона. При движении крови в ней постепенно возрастает осмотическая концентрация, а во время ее обратного движения к коре почки соли и другие растворенные вещества, диффундирующие через сосудистую стенку, переходят в интерстици-альную ткань. Тем самым сохраняется градиент концентрации осмотически активных веществ, т. е. прямые сосуды функционируют как противоточная система. Скорость движения крови по прямым сосудам влияет на количество удаляемых из мозгового вещества ионов Ма^ С1~ и мочевины, участвующих в создании осмотического градиента, и отток реабсорбируемой воды.
При водной нагрузке относительная проксимальная реабсорбция ионов и воды не изменяется, и в дистальный отдел нефрона поступает такое же количество жидкости, как и без нагрузки. При этом стенка дистальных отделов почечных канальцев остается водонепроницаемой, а из протекающей мочи клетки продолжают реабсорбировать соли натрия; при этом выделяется гипотоническая моча, концентрация осмотически активных веществ в которой ниже 50 мосмоль/л. Проницаемость канальцев для мочевины низкая, и она экскретируется с мочой, не накапливаясь в мозговом веществе почки. Собирательные трубки также обеспечивают реабсорбцию натрия, хлора и других ионов. Их основная функциональная особенность состоит в том, что реабсорбция веществ происходит в небольших количествах, но против наиболее значительного градиента, что обусловливает существенные различия концентрации ряда неорганических веществ в моче по сравнению с кровью. ,
Таким образом, деятельность петли нефрона, конечных частей дистального отдела собирательных трубок обусловливает способность почек человека при водной нарузке выделять большие объемы (до 900 мл/ч) разведенной, гипотонической мочи, а при дефиците воды в организме экскретировать мочи всего 10—12 мл/ч, в 4'/2 раза осмотически более концентрированной, чем кровь. Способность почки осмотически концентрировать мочу исключительно развита у некоторых пустынных грызунов, что позволяет им длительное время не пить воду.
ГОМЕОСТАТИЧЕСКАЯ ФУНКЦИЯ ПОЧЕК
Для поддержания почками постоянства объема и состава внутренней среды и прежде всего крови существуют специальные системы рефлекторной регуляции, включающие специфические рецепторы, афферентные пути и нервные центры, где происходит
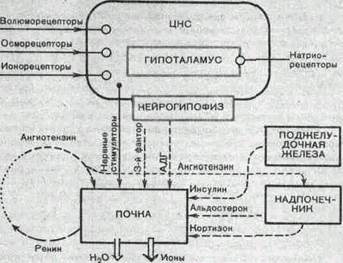
Рис.206. Схема регуляции водо- и электролитовыделительной функции почки.
переработка информации. Команды к почке поступают по эфферентным нервам или гуморальным путем. В целом перестройка работы почки, ее приспособление к непрестанно изменяющимся условиям определяется преимущественно влиянием на гломеру-лярный и канальцевый аппарат различных гормонов: АДГ, альдостерона, паратгормона и многих других (рис. 206)
Роль почек в осморегуляции и волюморегуляции. Почки являются основным органом осморегуляции. Они обеспечивают выделение избытка воды из организма в виде гипотонической мочи при увеличенном содержании воды (гипергидратация) или экономят воду и экскретируют мочу, гипертоническую по отношению к плазме крови при обезвоживании организма (дегидратация). При избыточном содержании воды в организме концентрация растворенных осмотически активных веществ в крови снижается и ее осмотическое давление падает. Это уменьшает активность центральных осморецепторов, расположенных в области супраоптического ядра гипоталамуса, а также периферических осморецепторов, имеющихся в печени, почке, селезенке и ряде других органов, что снижает выделение АДГ из нейрогипофиза в кровь и приводит к усилению выделения воды почкой.
При обезвоживании организма (или введении в сосудистое русло гипертонического раствора хлорида натрия) увеличивается концентрация осмотически активных веществ в плазме крови, возбуждаются осморецепторы, усиливается секреция АДГ, возрастает факультативная реабсорбция воды, уменьшается мочеотделение и выделяется осмотически концентрированная моча. В эксперименте на животных было показано, что секреция АДГ возрастает при раздражении не только осморецепторов, но и специфических натриорецепторов. Поэтому после введения в область III желудочка мозга гипертонического раствора хлорида натрия наступает антидиурез, а после введения в ту же область гипертонических растворов сахара угнетения мочеотделения не наблюдается.
Осморецепторы чутко реагируют на сдвиги концентрации осмотически активных веществ в плазме крови. При увеличении осмолярности плазмы крови на 1% концентрация'АДГ возрастает на 1 пг/мл (пикограмм равен 1 миллионной доле микрограмма). Переход же к состоянию максимального осмотического концентрирования мочи требует всего лишь 10-кратного возрастания количества АДГ в крови.
Помимо информации от осморецепторов и натриорецепторов, уровень секреции АДГ зависит от активности волюморецепторов, реагирующих на изменение объема
| 14* |
внутрисосудистой и внеклеточной жидкости. Ведущее значение в регуляции секреции АДГ имеют те волюморецепторы, которые реагируют на изменение напряжения сосудистой стенки в области низкого давления. В первую очередь следует назвать рецепторы левого предсердия, импульсы от которых передаются в ЦНС по афферентным волокнам блуждающего нерва. Изолированная активация волюморецепторов, возникающая в ответ на увеличение объема внутрисосудистой жидкости, приводит к повышению экскреции солей натрия и воды. Одновременная активация волюм- и осморецепторов в основном усиливает выведение воды, уменьшая ее реабсорбцию.
Роль почек в регуляции ионного состава крови. Почки являются важнейшим эффек-торным органом в системе ионного гомеостаза. Новейшие данные свидетельствуют о существовании в организме систем регуляции баланса каждого из ионов. Для некоторых из ионов уже описаны специфические рецепторы, например натриорецепторы. Появились первые данные о рефлекторной регуляции баланса других ионов.
Известны гормоны, регулирующие реабсорбцию и секрецию ионов в почечных канальцах. Реабсорбция натрия возрастает в конечных частях дистального отдела нефрона и собирательных трубках под влиянием гормона коры надпочечника альдосте-рона. Этот гормон выделяется в кровь при снижении концентрации натрия в плазме и уменьшении объема циркулирующей крови. Усиленное выделение натрия почкой вследствие угнетения его реабсорбции достигается при секреции в кровь так называемого натрийуретического гормона; его выработка возрастает при увеличении объема циркулирующей крови и повышении объема внеклеточной жидкости в организме. Хотя мысль о существовании этого гормона была высказана в конце 50-х годов, его структура и место секреции до сих пор не установлены.
Альдостерон усиливает выделение калия с мочой. Уменьшает выделение калия ин-сулин. Экскреция калия тесно связана с кислотно-щелочным состоянием. Алкалоз сопровождается усилением выделения калия с мочой, а ацидоз — его уменьшением.
При снижении концентрации кальция в крови паращитовидные железы выделяют паратгормон, который способствует нормализации уровня кальция в крови за счет увеличения его реабсорбции в почечных канальцах и повышения резсорбции кальция из костей. При гиперкальциемии под влиянием гастрина или подобного ему вещества, вырабатываемого в желудочно-кишечном тракте, стимулируется выделение в кровь клетками щитовидной железы гормона тирокальцитонина, который снижает концентрацию кальция в крови, способствует увеличению его экскреции почкой и переходу ионов Са'1"1' в кости.
В почечных канальцах регулируется также уровень реабсорбции магния, хлора, сульфатов и других ионов.
Роль почек в регуляции кислотно-основного состояния. Почки играют важную роль в поддержании постоянства концентрации ионов водорода в крови. Активная реакция мочи у человека и животных может очень резко изменяться, концентрации ионов водорода в моче при крайних состояниях работы почек различаются почти в 1000 раз (рН в некоторых случаях снижается до 4,5 или возрастает до 8,0, что и обеспечивает участие почек в стабилизации рН плазмы крови на уровне 7,36). Механизм закисления мочи основан на секреции клетками в просвет канальцев ионов водорода (рис. 207).
В апикальной плазматической мембране и цитоплазме клеток различных отделов нефрона находится фермент карбоангидраза, катализирующий реакцию гидратации двуокиси углерода.
сог+ню^Р^^-^га н.со.,,_н+ +1\со.^
Фильтрующийся из плазмы крови в просвет канальцев МаНСОз взаимодействует с ионами водорода, секретированными клеткой, приводя к образованию СОг. В просвете канальца ионы Н'1' связываются не только с НСОз, но и с такими соединениями, как двузамещенный фосфат (МагНРС^) и некоторыми другими, в результате чего увеличивается экскреция титруемых кислот с мочой. Это способствует восстановлению резерва
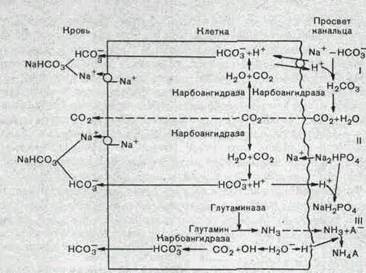
Рис.207. Схема основных реакций, обусловливающих участие почки в регуляции кислотно-основного равновесия. Объяснение в тексте.
оснований в плазме крови. Наконец, секретируемые ионы Н'1' могут связываться в просвете канальца с аммиаком, выделяющимся в клетке из глутамина и аминокислот (аммониогенез) и диффундирующим через мембрану в просвет канальца, в результате чего образуется ион аммония:
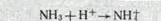
Таким образом, общая экскреция кислот почкой складывается из трех компонентов:
выделения НаСОз; титруемых кислот и выделения аммония МН^
Кислотовыделительная функция почек во многом зависит от состояния кислотно-основного состояния в организме.
При питании мясом образуется больше кислот и моча становится кислой, а при потреблении растительной пищи рН мочи сдвигается в щелочную сторону. При интенсивной физической работе из мышц в кровь поступают значительные количества молочной и фосфорной кислот. В этих условиях почки увеличивают выделение кислых продуктов. При гиповентиляции легких происходит задержка углекислого газа и снижается рН крови — развивается дыхательный ацидоз, при гипервентиляции уменьшается содержание углекислого газа в крови, растет рН крови — возникает состояние дыхательного алкалоза. Если в крови нарастает содержание ацетоуксусной и р-оксимасляной кислот, как это может наблюдаться при сахарном диабете, развивается состояние метаболического ацидоза. Рвота, сопровождающаяся потерей соляной кислоты, приводит к метаболическому алкалозу.
Легкие наряду с почками участвуют в нормализации кислотно-основного состояния. При дыхательном ацидозе увеличивается экскреция Н"1" и реабсорбция НСОз^ Метаболический ацидоз компенсируется гипервентиляцией легких.
В конечном счете почки, стабилизируя концентрацию ионов Н'1' в плазме крови, поддерживают рН на.уровне 7,36.
Экскреторная функция почек. Почки играют ведущую роль в выделении из крови нелетучих конечных продуктов обмена и чужеродных веществ, попавших во внутреннюю среду организма. В процессе метаболизма белков и нуклеиновых кислот образуются
различные продукты азотистого обмена. Основными азотсодержащими веществами мочи у человека являются мочевина, мочевая кислота, креатинин и др.
Катаболизм пуриновых оснований в организме человека останавливается на уровне образования мочевой кислоты (в клетках некоторых животных имеется полный набор ферментов, обеспечивающих распад пуриновых оснований до углекислоты и аммиака). Мочевая кислота в почке человека фильтруется в клубочках, затем реабсорбируется в каяальцах, часть мочевой кислоты секретируется клетками в просвет нефрона. Обычно у человека экскретируемая фракция мочевой кислоты довольно низкая—9,8%, что указывает на реабсорбцию значительных ее количеств в канальцах. В последние десятилетия непрерывно возрастает интерес к изучению механизмов транспорта мочевой кислоты в почечных канальцах в связи с резко возросшей частотой заболевания подагрой, при которой нарушен обмен мочевой кислоты.
Весь образующийся в течение суток креатинин, источником которого служит креа-тинфосфорная кислота, выделяется почками с мочой. Его суточная продукция и экскреция зависят не столько от потребления мяса с пищей, сколько от массы мышц тела. Креатинин, как и мочевина, свободно фильтруется в почечных клубочках, но с мочой выводится весь профильтровавшийся креатинин, в то время как мочевина частично реабсорбируется в канальцах.
Помимо перечисленных имеется еще очень много разнообразных веществ, непрестанно удаляемых почкой из крови. О том, какие вещества удаляет или разрушает почка, можно судить при изучении состава крови у людей с удаленными почками. В их крови, помимо мочевины, креатинина, мочевой кислоты, накапливаются гормоны (глюкагон, паратгормон, гастрин), ферменты (рибонуклеаза, ренин), производные индола, глюку-роновая кислота и многие другие вещества.
Необходимо помнить, что даже физиологически ценные вещества при их избытке в крови начинают экскретироваться почкой. Это относится как к неорганическим веществам, о которых шла речь выше при описании осмо-, волюм- и ионорегулирующей функции почек, так и органическим веществам, таким, как глюкоза, аминокислоты. Повышенная экскреция этих веществ может в условиях патологии наблюдаться и при нормальной их концентрации в крови, когда нарушена работа клеток, реабсорбирующих то или иное профильтровавшееся вещество из канальцевой жидкости в кровь.
Инкреторная функция почек. В почке вырабатывается несколько физиологически активных веществ, позволяющих рассматривать ее как важный инкреторный орган. Гранулярные клетки юкстагломерулярного аппарата под влиянием ряда стимулов (при уменьшении артериального давления в почке, снижении содержания натрия в организме и при переходе из горизонтального в вертикальное положение) секретируют ренин. Ренин, синтезируемый в гранулярных клетках, представляет собой протеолитический фермент. В плазме крови он отщепляет от «г-глобулина (ангиотензиноген) физиологически неактивный пептид, состоящий из 10 аминокислот — ангиотензин I. Затем от анги-отензина I отщепляются две аминокислоты и он превращается в очень активное сосудосуживающее вещество — ангиотензин II. Помимо влияния на сосуды и тем самым на величину артериального давления, ангиотензин II, в частности, усиливает секрецию альдостерона, усиливает чувство жажды, регулирует реабсорбцию натрия в канальцах.
Клетки почки извлекают из плазмы крови образующийся в печени прогормон — витамин Оз и превращают его в физиологически очень активный гормон. Этот стероид стимулирует образование кальцийсвязывающего белка в кишечнике, способствует освобождению кальция из костей, регулирует его реабсорбцию в почечных канальцах.
В почке синтезируется активатор плазминогена — урокиназа. В мозговом веществе почки образуются простагландины, в том числе простагландин Аа, который называют также медуллином. Они участвуют в регуляции общего и почечного кровотока, вызывают натрийурез, уменьшают чувствительность клеток к АДГ. В почке образуется эри-трогенин, который способствует превращению в плазме крови неактивного эритропоэ-тиногена в эритропоэтин, стимулирующий эритропоэз в костном мозге. В почке вырабатывается брадикинин, являющийся сильным вазодилататором.
Метаболическая функция почек. Важной стороной работы почек является их участие в обмене белков, углеводов и липидов. Не следует смешивать понятия метаболизма самих почек (т. е. процессы обмена веществ в почке, обеспечивающие все многообразие выполняемых ею функций) и метаболическую функцию почек, связанную с их участием в обеспечении постоянства содержания в крови ряда органических веществ. В почечных клубочках фильтруются низкомолекулярные белки, пептиды. Клетки проксимального ка-нальца нефрона расщепляют их до аминокислот, которые через базальную плазматическую мембрану транспортируются во внеклеточную жидкость и затем в кровь. Это способствует восстановлению в организме фонда аминокислот. При заболеваниях почки эта функция может нарушаться.
В почке имеется система новообразования глюкозы — глюконеогенез. Так, при длительном голодании почки могут синтезировать половину от общего количества глюкозы, поступающей в кровь. В почке синтезируется фосфатидилинозит, являющийся необходимым компонентом плазматических мембран. Значение почки в липидном обмене состоит в том, что свободные жирные кислоты в ее ткани могут включаться в состав три-ацилглицерина и фосфолипидов и в виде этих соединений могут поступать в кровь.
Дата добавления: 2016-03-27; просмотров: 2818;