ЗРИТЕЛЬНЫЕ ПИГМЕНТЫ. ФОТОРЕЦЕПЦИЯ
В палочках сетчатки человека содержится пигмент родопсин, или зрительный пурпур, максимум спектра поглощения которого находится в области 500 нанометров (нм). В наружных сегментах трех типов колбочек (сине-, зелено- и красно-чувствительных) содержится три типа зрительных пигментов, максимумы спектров поглощения которых находятся в синей (420 нм), зеленой (531 нм) и красной (558 нм) частях спектра. Красный колбочковый пигмент получил название «йодопсин» (поглощает желтую часть спектра). Молекула зрительного пигмента сравнительно небольшая, состоит из большей белковой части (опсина) и меньшей хромофорной (ретиналь, или альдегид витамина А). Ретиналь может находиться в различных пространственных конфигурациях, т. е. изомерных формах, но только одна из них — 11-цис-изомер ретиналя выступает в качестве хромофорной группы всех известных зрительных пигментов. Источником ретиналя в организме служат каротиноиды, поэтому недостаток их приводит к дефициту витамина А и, как следствие, к недостаточному ресинтезу родопсина, что в свою очередь является причиной нарушения сумеречного зрения, или «куриной слепоты».
Молекулярная физиология фоторецепции.
|
| ||||
| Рис. 14. Изомерные формы молекулы зрительного пигмента: А –в темноте, Б –на свету |
В темноте ретиналь в виде цис-формы (рис. 14 А). На свету меняет свою конфигурацию и превращается в транс-форму (рис. 14 Б). Его боковая цепь выпрямляется. Связь ретиналя и белка прерывается. Разложение пигмента сопровождается его выцветанием, при этом высвобождается энергия, которая создает ПД, который через синапс запускает импульс в нейронах. Обратное превращение пигмента родопсина происходит при затемнении глаз. Для образования ретиналя необходим цис-изомер витамина А. Если витамин А в организме отсутствует, развивается куриная слепота (человек не видит в сумерках).
Опсин при воздействии кванта света тоже меняется. Происходит перемещение заряда на белке. Этот процесс ведет к возникновению раннего рецепторного потенциала (РРП). Вслед за РРП развивается поздний РРП, который отражает возбуждение нервного членика рецептора – внутреннего сегмента. ПРП через синапс запускает импульс в нейронах. Структура йодопсина близка к родопсину (тоже состоит из ретиналя с белком опсином).
НЕЙРОНЫ СЕТЧАТКИ
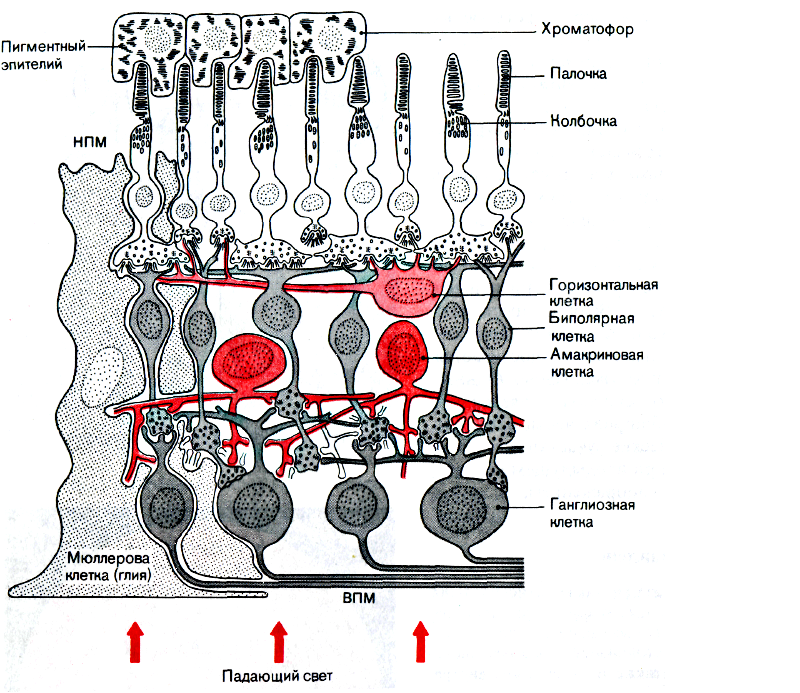
|
| Рис. 15. Схема строения сетчатки (по данным электронной микроскопии). НПМ-наружная пограничная мембрана, ВПМ-внутренняя пограничная мембрана |
Фоторецепторы сетчатки синаптически связаны с биполярными нейронами. При действии света уменьшается выделение медиатора (глутамата) из фоторецептора, что приводит к гиперполяризации мембраны биполярного нейрона. От него нервный сигнал передается на ганглиозные клетки, аксоны которых являются волокнами зрительного нерва. Передача сигнала как с фоторецептора на биполярный нейрон, так и от него на ганглиозную клетку происходит безымпульсным путем. Биполярный нейрон не генерирует импульсов ввиду предельно малого расстояния, на которое он передает сигнал.
На 130 млн фоторецепторных клеток приходится только 1 млн 250 тыс. ганглиозных клеток, аксоны которых образуют зрительный нерв. Это значит, что импульсы от многих фоторецепторов сходятся (конвергируют) через биполярные нейроны к одной ганг-лиозной клетке. Фоторецепторы, соединенные с одной ганглиозной клеткой, образуют рецептивное поле ганглиозной клетки. Рецептивные поля различных ганглиозных клеток частично перекрывают друг друга. Таким образом, каждая ганглиозная клетка суммирует возбуждение, возникающее в большом числе фоторецепторов. Это повышает световую чувствительность, но ухудшает пространственное разрешение. Лишь в центре сетчатки, в районе центральной ямки, каждая колбочка соединена с одной так называемой карликовой биполярной клеткой, с которой соединена также всего одна ганглиозная клетка. Это обеспечивает здесь высокое пространственное разрешение, но резко уменьшает световую чувствительность.
Взаимодействие соседних нейронов сетчатки обеспечивается горизонтальными и амакриновыми клетками, через отростки которых распространяются сигналы, меняющие синаптическую передачу между фоторецепторами и биполярными клетками (горизонтальные клетки) и между биполярными и ганглиозными клетками (амакриновые клетки). Амакриновые клетки осуществляют боковое торможение между соседними ганглиозными клетками (рис. 15).
Кроме афферентных волокон, в зрительном нерве есть и центробежные, или эфферентные, нервные волокна, приносящие к сетчатке сигналы из мозга. Полагают, что эти импульсы действуют на синапсы между биполярными и ганлиозными клетками сетчатки, регулируя проведение возбуждения между ними.
29. СВЕТОВАЯ И ТЕМНОВАЯ АДАПТАЦИЯ
При переходе от темноты к свету наступает временное ослепление, а затем чувствительность глаза постепенно снижается. Это приспособление зрительной сенсорной системы к условиям яркой освещенности называется световой адаптацией. Обратное явление (темповая адаптация) наблюдается при переходе из светлого помещения в почти не освещенное. В первое время человек почти ничего не видит из-за пониженной возбудимости фоторецепторов и зрительных нейронов. Постепенно начинают выявляться контуры предметов, а затем различаются и их детали, так как чувствительность фоторецепторов и зрительных нейронов в темноте постепенно повышается.
Повышение световой чувствительности во время пребывания в темноте происходит неравномерно: в первые 10 мин она увеличивается в десятки раз, а затем в течение часа — в десятки тысяч раз. Важную роль в этом процессе играет восстановление зрительных пигментов. Пигменты колбочек в темноте восстанавливаются быстрее родопсина палочек, поэтому в первые минуты пребывания в темноте адаптация обусловлена процессами в колбочках. Этот первый период адаптации не приводит к большим изменениям чувствительности глаза, так как абсолютная чувствительность колбочкового аппарата невелика.
Следующий период адаптации обусловлен восстановлением родопсина палочек. Этот период завершается только к концу первого часа пребывания в темноте. Восстановление родопсина сопровождается резким (в 100 000—200 000 раз) повышением чувствительности палочек к свету. В связи с максимальной чувствительностью в темноте только палочек слабо освещенный предмет виден лишь периферическим зрением.
Существенную роль в адаптации, помимо зрительных пигментов, играет изменение (переключение) связей между элементами сетчатки. В темноте площадь возбудительного центра рецептивного поля ганглиозной клетки увеличивается вследствие ослабления или снятия горизонтального торможения. При этом увеличивается конвергенция фоторецепторов на биполярные нейроны и биполярных нейронов на ганглиозную клетку. Вследствие этого за счет пространственной суммации на периферии сетчатки световая чувствительность в темноте возрастает.
Световая чувствительность глаза зависит и от влияний ЦНС. Раздражение некоторых участков ретикулярной формации ствола мозга повышает частоту импульсов в волокнах зрительного нерва. Влияние ЦНС на адаптацию сетчатки к свету проявляется и в том, что освещение одного глаза понижает световую чувствительность неосвещенного глаза. На чувствительность к свету оказывают влияние также звуковые, обонятельные и вкусовые сигналы.
Дата добавления: 2015-11-28; просмотров: 2255;