Р. Шмидт
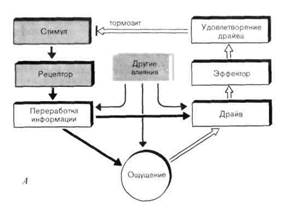
Чувство жажды, которое мы испытываем, когда не выпили достаточно жидкости, и чувство голода-когда давно не ели, нельзя отнести к какому-то конкретному органу или части тела. Поэтому они называются «общими чувствами». Другими примерами общих чувств являются усталость, ощущение духоты и половое влечение. С точки зрения сенсорной физиологии все они характеризуются тем, что вызываются одним или несколькими адекватными стимулами, возникающими в самом организме, а не в окружающей среде. Эти стимулы обнаруживаются рецепторами (часть из которых еще неизвестна) и вызывают соответствующие общие ощущения (рис. 9-1, А). Например, ниже мы расскажем, как «сгущение» жидкости в организме из-за недостатка воды воспринимается осморецепторами, и в результате возникает чувство жажды (рис. 9-1, Б). Можно представить себе также, что в течение дня в крови накапливаются конечные продукты обмена и они вызывают чувство усталости, или же некоторые гормоны вызывают половое влечение или благоприятствуют его появлению, когда их концентрация в организме достаточно высока.
Стимулы, способные вызывать общие ощущения, вызывают также разные виды активности, направленные на снижение таких ощущений или их устранение. Иными словами, стимулы, вызывающие общие ощущения, приводят к драйвам (drive-побуждение)-мотивационным состояниям, которые побуждают организм добывать то, чего ему недостает. Хотя такие драйвы контролируются ощущениями, они могут возникать также и независимо (рис. 9-1, А). Недостаточное количество воды в организме ведет не только к ощущению жажды, но и к поискам воды, и если эти поиски успешны, то недостаток ее устраняется (рис. 9-1, Б). В самом общем виде это значит, что удовлетворение побуждения устраняет причину, вызвавшую общее чувство.
Драйвы, связанные с общими чувствами, способствуют выживанию индивидуума или вида. Поэтому, как правило, они должны удовлетворяться. Это врожденные состояния, не требующие обучения. Но в течение жизни многочисленные влияния видоизменяют их, в особенности на высших филогенетических уровнях. Эти влияния действуют в разные моменты всего процесса. Описание драйвов и их изменений не входит в задачу настоящей книги; здесь будут рассмотрены как примеры общих чувств только те аспекты жажды и голода, которые связаны с сенсорной физиологией.
Жажда
Условия возникновения чувства жажды.Организм взрослого человека на 70-85% (по весу) состоит из воды (не считая отложений жира). Это высокое содержание воды колеблется в очень узких пределах. В норме оно колеблется только на ± 0,22% веса тела, так что у человека весом 70 кг колебания эти составляют +150 мл. При потере воды больше
|
 Рис. 9-1. Отношения между общими чувствами и драйвами. А. Общий характер взаимоотношений между ощущениями и драйвами. Б. Возникновение чувства жажды и активация поиска воды при ее нехватке. В ощущении жажды помимо осморецепторов участвуют и другие рецепторы (см. рис. 9-2). Вертикальные красные полоски на кончиках верхних стрелок означают, что удовлетворение драйва ведет к устранению стимула.
Рис. 9-1. Отношения между общими чувствами и драйвами. А. Общий характер взаимоотношений между ощущениями и драйвами. Б. Возникновение чувства жажды и активация поиска воды при ее нехватке. В ощущении жажды помимо осморецепторов участвуют и другие рецепторы (см. рис. 9-2). Вертикальные красные полоски на кончиках верхних стрелок означают, что удовлетворение драйва ведет к устранению стимула.
0,5% веса тела (около 350 мл у человека весом 70 кг) возникает жажда. Физиологически организм может терять воду четырьмя путями: с мочой, с потом, в виде водяных паров в выдыхаемом воздухе и (обычно в ограниченном количестве) с калом. Эта вода должна быть возмещена, чтобы не нарушилось жизненно важное тонко сбалансированное равновесие между множеством растворенных веществ. Как показывают данные по водному балансу (табл. 9-1), в умеренном климате человек, потребляющий обыкновенную пищу, получает воду главным образом в виде питья, а также как составную часть твердой пищи и в меньшей степени-как продукт окислительного распада этой пищи в организме. Суммарные цифры выражают суточный кругооборот воды у взрослого человека, составляющий около 3-4% веса тела.
Еще одним фактором, влияющим на чувство жажды, является распределение воды в организме на две части (компартмента) с разными свойствами. Около двух третей жидкости тела содержится в клетках, во внутриклеточном.. пространстве, а одна треть во внеклеточном пространстве. Около трех четвертей внеклеточной воды находится в промежутках между клетками, в интерстициальном пространстве, а остальная - в сосудистой системе,
Таблица 9-1. Суточный водный баланс у взрослого человека (Muntwyler, 1973)
| Поступление воды мл | Потери воды мл |
| Питье 1200 | С мочой 1400 |
| С твердой пищей 900 | Через легкие и кожу 900 |
| В процессе метаболизма 300 | С калом 100 |
| Всего 2400 |
где она образует водную фазу плазмы крови (крови без кровяных телец). Вне- и внутриклеточные жидкости разделены клеточными мембранами и имеют разный состав, в особенности в отношении катионов и анионов. В интересующем нас аспекте главное различие состоит в том, что внеклеточная жидкость содержит большие количества Na+, а внутриклеточная богата К+, но содержит мало Na+. Плазма крови и интерстициальная жидкость сообщаются через стенки капилляров; они почти не различаются по содержанию солей, но белка в плазме значительно больше, чем в интерстициальной жидкости.
Клеточные мембраны, разделяющие внутри- и внеклеточные пространства, легко проницаемы для воды, но для солей их проницаемость значительно ниже. Для простоты изложения мы примем, что мембраны непроницаемы для солей. В конечном счете диффузия воды через такие полупроницаемые мембраны (т. е. проницаемые только для воды) всегда направлена к пространству с более высокой концентрацией солей (т.е. с меньшей «концентрированностью воды»). При равной концентрации соли в обоих пространствах вода диффундирует сквозь клеточную мембрану одинаково в обе стороны, в итоге перемещения не происходит. Нo если из одного пространства удалить воду, так что концентрация соли в нем повысится, то вода будет перемещаться сюда из второго пространства, пока не выравняются концентрации. Этот процесс называется осмосом, а разница в гидростатическом давлении, которая точно противодействовала бы стремлению воды диффундировать через полупроницаемую мембрану между пространствами, содержащими жидкости с разной концентрацией солей, называется осмотическим давлением. Солевой раствор с таким осмотическим давлением, как у нормальной жидкости тела, называется изотоническим. Отсюда гипертоническим и гипотоническим называют растворы солей соответственно более и менее концентрированные, чем нормальная тканевая жидкость. Например, если вода удаляется из плазмы вследствие образования мочи в почке, то произойдет диффузия воды из интерстициального пространства в плазму, а из клеток в интерстициальное пространство. Если же клетка в потовой железе секретирует пот (т.е. теряет воду), то вода должна диффундировать из интерстициального пространства в клетку железы. • Физиологические потери воды организмом (с мочой, потом, влагой в выдыхаемом воздухе) все вместе приводят к потере воды как из внеклеточного, так и из внутриклеточного пространства. Одновременно повышается концентрация солей в этих растворах; они становятся гипертоническими, но обычно лишь в незначительной степени. Дальнейшим следствием является уменьшение секреции слюны, что приводит к ощущению сухости во рту и в глотке, столь характерному для жажды. Если бы существовали соответствующие рецепторы, можно было бы измерить потерю воды организмом а) по объему или осмотическому давлению клеток, б) по объему или осмотическому давлению внеклеточного пространства или в) косвенно по снижению секреции слюны и возникающей при этом сухости слизистых оболочек.
Адекватные стимулы для чувства жажды.Чтобы решить, которое из перечисленных изменений может быть причиной чувства жажды, каждый из этих факторов нужно изучить экспериментально по отдельности; иначе говоря, содержание воды и солей в одном пространстве надо менять, не затрагивая второго, или же надо менять одну только секрецию слюны. Такие опыты проведены на животных, причем количество выпиваемой воды служило показателем степени экспериментально вызванной жажды. Наиболее важные результаты этих опытов изложены ниже.
После внутривенного вливания гипертонического раствора собака выпивает воды вдвое больше, чем после внутривенного вливания осмотически эквивалентного раствора мочевины. В первом случае из-за непроницаемости клеточных мембран для ионов Na+ вода выходит из клеток. Но для мочевины клеточные мембраны легко проницаемы; поэтому, когда ее вводят, концентрации во внутри- и внеклеточных пространствах выравниваются при заметно меньшем изменении объема клеток и их тоничности (концентрации соли). Этот факт подтвердился в повторных опытах со многими модификациями на разнообразных  млекопитающих. Отсюда следует вывод, что уменьшение объёма клеток без изменения со дерзания в них солей (но не концентрации, которая возрастает)вызывает жажду.
млекопитающих. Отсюда следует вывод, что уменьшение объёма клеток без изменения со дерзания в них солей (но не концентрации, которая возрастает)вызывает жажду.
Если количество Na+ во внеклеточном пространстве снизить в опыте (например, изменив состав пищи или посредством искусственной почки), то оно теряет воду; часть ее выходит из тела, а часть диффундирует в клетки. В этих условиях, даже несмотря на то что объем клеток возрастает, появляется жажда. (Возникает также потребность в соли, но этого мы здесь касаться не будем). Кроме того, можно экспериментально снизить объем внеклеточной жидкости, не меняя концентрацию NaCl в ней, так что объем клеток не меняется; в этом случае также возникает жажда. Отсюда можно сделать вывод, что уменьшение объема внеклеточной жидкости тоже вызывает жажду. Опыты показали, что эффекты этих двух факторов аддитивны; при одновременном понижении объема клеток и объема внеклеточной жидкости жажда становится особенно сильной.
Сухость во рту, сопровождающая все формы жажды, вызывается, как уже было указано, снижением секреции слюны. Это явление отражает недостаток воды. Но в отличие от прежних толкований оно, по-видимому, представляет собой не причину, а скорее симптом, связанный с чувством жажды. Об этом говорят следующие данные. Смачивание-поверхности рта_и_ глотки не__устраняет жажды, хотя может немного облегчить ее. Жажду также не ослабляет и не предотвращает местная анестезия слизистой рта или даже полная денервация области рта и глотки. Врожденное отсутствие слюнных желез (у людей) или их оперативное удаление (у животных) не оказывает заметного действия на потребление воды; количество выпиваемой воды остается нормальным. Наконец, внутривенное внедрение воды немедленно приносит облегчение, хотя сухость во рту и глотке какое-то время может сохраняться.
Рецепторы и центральные механизмы.Нервные структуры, преимущественно регулирующие водно-солевой баланс, находятся в промежуточном мозгу (диэнцефалон), в основном в гипоталамусе и около него. Эта часть головного мозга имеет прямое отношение к регуляции вегетативных функций. Опытами здесь выявлено наличие осморецепторов, главным образом в областях- впереди гипоталамуса. Это сенсорные элементы, реагирующие на повышение концентрации соли внутри клеток при потере ими воды. Например, после введения козе очень малых количеств (меньше 0,2 мл) гипертонического раствора NaCl в некоторые части этой области животное через 30-60 с начинало пить и в течение 2-5 мин поглощало 2-8 л воды. Электрическая стимуляция этих же нервных структур тоже заставляет животных много пить. Хирургическое удаление или коагуляция некоторых участков гипоталамуса может ослабить или совсем прекратить питье даже при нехватке воды в организме. Все эти данные показывают, что осморецепторы в промежуточном мозгу служат сенсорными аппаратами жажды, вызываемой дефицитом воды в клетках (рис. 9-2). Нейронные структуры в гипоталамусе несомненно играют решающую роль в переработке информации от этих осморецепторов.
Что касается сенсорных элементов, на которых основано чувство жажды, вызываемое дефицитом воды во внеклеточном пространстве, то здесь имеются только предположения и косвенные данные. В настоящее / время представляется вероятным, что рецепторы растяжения в стенках крупных вен и в предсердиях не только влияют на кровообращение, но также участвуют в регуляции водного баланса и возникновения жажды (рис. 9-2). Гипоталамус служит важным центром переработки сигналов, приходящих от рецепторов растяжения (по афферентным волокнам блуждающего нерва). Внеклеточная дегидратация связана также с секрецией ренина и образованием ангиотензина 11.Ангиотензин 11 – вещество, весьма действенно вызывающее жажду при введении его в кровоток или непосредственно в разные отделы гипоталамуса, в том числе субфорникальный орган. Ангиотензин несомненно способствует возникновению жажды, связанной с уменьшением объема жидкости (гиповолемия), но пока что относительное значение ангиотензина и других механизмов для такого рода жажды еще не определено.

Рис. 9-2. Возникновение чувства жажды. Участвующие в этом рецепторы показаны серым. Прямоугольники над ними означают их адекватные стимулы. Сухость во рту является косвенным следствием внутриклеточного и внеклеточного дефицита вода.
Об уменьшении слюноотделения, связанном с дефицитом воды и приводящем к сухости во рту, сигнализуют рецепторы в слизистых оболочках рта и глотки. В опытах на животных обнаружены здесь разные виды рецепторов - механорецепторы, холодовые и тепловые рецепторы и, возможно, водяные рецепторы. Неизвестно, в какой степени каждый из них участвует в создании этого периферического компонента жажды. Если эти рецепторы стимулируются, когда в организме нет общего недостатка воды, что возможно, когда человек говорит, курит, дышит через рот или ест очень сухую пищу, то вызываемую ими «ложную жажду» можно утолить, смочив слизистую рта. Но сигналы, идущие из области рта и глотки, совсем не обязательны, если чувство жажды вызывается действительным дефицитом воды в организме. При истинной жажде, как указано выше, смачивание глотки может уменьшить, но не утолить жажду.
Таким образом, жажда - это общее ощущение, основанное на интегрированной реакции многих типов рецепторов, из которых одни находятся на периферии, а другие в ЦНС. Они приведены на рис. 9-2. Промежуточный мозг, и в частности гипоталамус, по-видимому, играет доминирующую роль в интеграции этого множества афферентных входов. Мы не знаем, насколько точно результаты опытов на животных приложимы к человеку, и также не знаем, какие центральные структуры вызывают чувство жажды. Но можно думать, что отношения, изображенные на рис. 9-2, распространяются и на человека.
Для чувства жажды не наступает адаптации. Этот субъективный опыт опять-таки подтверждается экспериментами на животных. Показано, что количество воды, потребляемое после внутривенного вливания гипертонического раствора соли, не зависит от быстроты вливания. Иными словами, жажда, вызываемая введением определенного количества раствора, оставалась неизменной независимо от того, возрастала концентрация NaCl очень медленно или очень быстро. Поскольку адаптация к жажде не наступает, единственный способ утолить ее-это выпить воды (рис. 9-1, Б).
Утоление жажды. Между моментом начала питья и устранением дефицита воды проходит значительное время; вода в желудке и кишечнике сначала должна всосаться (т.е. перейти в кровоток), что происходит главным образом в тонком кишечнике. Но как известно из обычных наблюдений и как неоднократно подтверждали опыты на животных, чувство жажды исчезает (т.е. питье прекращается) задолго до того, как дефицит воды во вне- и внутриклеточном пространствах мог быть возмещен. Это значит, что утолению жажды, наступающему после всасывания (absorption) воды,-так называемому постабсорбтивному -предшествует преабсорбтивное утоление; таким образом предотвращается потребление чрезмерного количества воды за период, пока усвоенная вода не окажет своего действия (рис. 9-3). Опыты на животных показывают, что этот преабсорбтивный механизм действует с большой точностью: количество выпитой воды очень близко соответствует тому, какое требуется в конечном счете.
Рецепторы и механизмы, лежащие в основе преабсорбтивного утоления жажды, неизвестны. Собака с фистулой в пищеводе пьет почти вдвое больше воды, чем нормальная собака с таким же дефицитом воды, а затем перестает пить на 20-60 мин. Следовательно, само по себе питье или связанные с ним моторные и сенсорные процессы вызывают некоторое временное утоление жажды. Растяжение желудка проглоченной жидкостью, по-видимому, тоже имеет значение. Вода, введенная крысам и другим животным непосредственно в желудок, соответственно снижает количество выпиваемой воды. Но связанные с этим нервные механизмы остаются неясными.
 Рис. 9-3. Утоление жажды (преабсорбтивное и постабсорбтивное). Рецепторы, показанные на рис. 9-2, представлены здесь одним прямоугольником (рецепторы жажды).
Рис. 9-3. Утоление жажды (преабсорбтивное и постабсорбтивное). Рецепторы, показанные на рис. 9-2, представлены здесь одним прямоугольником (рецепторы жажды).
После того как дефицит воды действительно компенсирован (постабсорбтивное утоление жажды), проходит некоторое время, пока чувство жажды возобновится, даже если происходит непрерывная (хотя и медленная) физиологическая потеря воды. Это значит, что жажда имеет свой порог. Как уже было сказано выше, этот порог эквивалентен потере воды, приблизительно равной 0,5% веса тела. Порог ограничивает появление жажды определенными интервалами; без этого неэффективно уходило бы много времени на питье малых количеств воды. Итак, содержание воды в организме человека колеблется между максимумом после постабсорбтивного утоления жажды и минимумом, который в идеальном случае лежит немного ниже порога. Но нередко колебания в содержании воды в организме выходят за эти пределы. Одна из причин этого состоит в том, что часто мы потребляем воды больше, чем нужно для возмещения ее потерь; другая причина та, что не всегда есть возможность утолить жажду сразу же, как только она становится заметной.
Первичное и вторичное питье.Питье, вызываемое абсолютным или относительным дефицитом воды в одном из пространств тела, содержащих жидкость, называется первичным питьем, а питье без видимой нужды в восполнении потерянной воды -вторичным питьем. Собственно говоря, вторичное питье служит обычным путем поступления воды. Обычно мы (и другие млекопитающие) выпиваем физиологически потребную воду заранее. Например, жидкость пьют за едой и после еды-очевидно, мы научились приноравливать выпиваемое количество к характеру пищи. Когда еда соленая, мы пьем больше, даже не ощущая жажды. Известную роль, по-видимому, играет также привычка, но мы располагаем лишь весьма скудными сведениями о механизмах, посредством которых потребность в воде оценивается заранее. Во всяком случае, первичное питье -это реакция, осуществляемая в особых случаях, и к нему редко прибегают люди, ведущие размеренный образ жизни.
Патологическая жажда.Усиление жажды во время болезни может быть следствием ненормально большой потери воды без нарушения механизмов жажды. Но оно может также означать нарушение этих механизмов или регуляции водно-солевого баланса. Ярким примером первого случая служит потеря воды, связанная с длительной рвотой или сильным поносом, как это бывает, например, при холере. Еще одним хорошо известным примером того же порядка является несахарный диабет, при котором из-за недостатка антидиуретического гормона (АДГ) из организма выводится ежесуточно много литров гипотонической мочи. Такие больные страдают от неутолимой жажды, и все дневные дела связаны у них с постоянной необходимостью пить. Детали многих сторон патологической жажды можно найти в руководствах по патофизиологии и клинической медицине.
В 9.1. Вода в организме содержится в двух больших пространствах-внутриклеточном и внеклеточном. Отношение внутриклеточного объема к внеклеточному составляет
а) 5:1;
б) 2:1;
в)1׃1
г) 1:3;
д) 1 :10.
В 9.2. Тело человека состоит на 70-75% (по весу) из воды. Потеря какой части веса тела является порогом для появления чувства жажды''
а) 0,05%.
б) 0,1%.
г)0,5%.
г) 1%.
Д) 3%.
В 9.3. Какие факторы участвуют в возникновении жажды? а)_повышение тоничности внутриклеточной жидкости.
б) Понижение тоничности внутриклеточной жидкости.
в) Увеличение объема клетки.
г)_Уменьшение объема внеклеточной жидкости.
д) Понижение тоничности внеклеточной жидкости.
В 9.4. Какие рецепторы сигнализируют о дефиците воды в организме?
а) Хеморецепторы в каротидной железе.
Б )__Осморецепторы в промежуточном мозгу.
В) Рецепторы растяжения в желудке.
Г) Рецепторы растяжения мышц, участвующих в акте глотания.
д) Рецепторы растяжения в крупных венах.
В 9.5. Которая из следующих форм питья чаще возмещает потери жидкости у человека?
а) Первичное питье.
_б)_Вторичное питье.
в) Первичное и вторичное питье используются одинаково часто.
Голод
Потребность в пище.Энергетический баланс сохраняется у любого животного, если содержание энергии в пище соответствует энергии, расходуемой при мышечной работе, в метаболических процессах (рост, перестройка) и при теплопотерях. Чрезмерное потребление пищи приводит к отложению жира и тем самым к увеличению веса, а недостаточное количество пищи ведет сначала к мобилизации и использованию отложений жира, а при длительном недоедании нарушает функции организма и иногда кончается смертью.
Человек и животные в норме приноравливают количество потребляемой пищи к меняющимся потребностям в зависимости от производимой работы, климата и питательной ценности пищи. Кратковременная регуляция принятия пищи накладывается на долговременную регуляцию, которая возмещает временные неправильности питания и обеспечивает возвращение к нормальному весу тела. Например, когда животные жиреют из-за насильственного кормления, а затем их возвращают в нормальные условия, они едят меньше, чем контрольные. Когда же они восстановят свой истинный контрольный вес, поедание пищи постепенно увеличивается. И наоборот, после периода голодания первоначальный вес тела приобретается вновь путем временного усиленного потребления пищи.
Heдостаток пищи вызывает чувство голода, и связанный с ним пищевой драйв ведет к приему пищи и в конечном счете к сытости (рис. 9-1, А). Какие механизмы вызывают голод и чувство насыщения? Возникает также вопрос, лежат ли в основе кратковременной и долговременной регуляции потребления пищи одни и те же или разные механизмы? Несмотря на многочисленные исследования, на эти вопросы еще не получены исчерпывающие ответы. Пробелы в наших знаниях станут очевидными из дальнейшего изложения.
Одно установлено с очевидностью, а именно что в ощущениях голода и сытости участвует несколько факторов. Но совершенно неизвестно, каково их относительное значение, а также неясно, все ли относящиеся сюда факторы уже обнаружены.
Факторы, вызывающие голод. Субъективный опыт показывает, что голод представляет собой общее, чувство локализующееся в области желудка (или проецирующееся на эту область); оно возникает, когда желудок пустой, и исчезает или уступает место чувству насыщения, как только желудок наполняется пищей. Некоторые прежние исследователи этого вопроса считали, что чувство голода вызывается сокращениями пустого желудка. По мнению этих авторов, такое представление согласуется с тем фактом, что кроме обычных сокращений, посредством которых пища перерабатывается и перемещается, сокращается также и пустой желудок. Такие сокращения, по-видимому, тесно связаны с голодом и поэтому могли бы способствовать возникновению этого чувства. Возможно, что о них сигнализируют в ЦНС механорецепторы в стенке желудка (рис. 9-4, слева). Но действие сокращений пустого желудка на голод не стоит переоценивать; при денервации желудка или полном его удалении в опыте на животных их пищевое поведение практически не меняется. Следовательно, такие сокращения могут быть одним из факторов, приводящих к ощущению голода, но фактором не обязательным.
Решающую роль в возникновении ощущения голода, по-видимому, играет глюкоза (виноградный сахар). Этот сахар служит главным источником энергии для клеток организма. Уровень глюкозы в крови и доступность глюкозы для отдельных клеток контролируются гормонами. Экспериментально показано, что снижение доступности глюкозы (не сам по себе уровень сахара в крови) очень хорошо коррелирует с чувством голода и мощными сокращениями желудка, т. е. фактор «наличия глюкозы» является решающим параметром в развитии голода.
Эта «глюкостатическая» гипотеза подтверждается различными экспериментальными данными, показывающими, что в промежуточном мозгу, печени, желудке и тонком кишечнике существуют глюкорецеп-торы. Например, при инъекции мышам золото-тиоглюкозы (золото_ является ядом для клеток) многие клетки в промежуточном мозгу, которые, очевидно, поглощают особенно большие количества глюкозы, разрушаются; при этом резко нарушается пищевое поведение. Иными словами, глюкорецепторы обычно сигнализируют о снижении количества наличной глюкозы и таким образом вызывают голод (рис. 9-4).
Выдвинуто и другое представление о том, каким путем вызывается голод, но в его пользу имеется меньше экспериментальных данных, чем для глюкостатической гипотезы. Это термостатическая гипотеза, основанная на наблюдении, что теплокровные животные поедают пищу в количествах, обратно пропорциональных температуре среды. Чем ниже температура окружающей среды, тем больше они едят, и наоборот. Согласно этой гипотезе, датчиками в процессе интеграции общего энергетического баланса служат внутренние (центральные) терморецепторы (см. разд. 3.3). В таком случае снижение общей теплопродукции влияет на внутренние терморецепторы, которые вызывают чувство голода (рис. 9-4). Можно показать экспериментально, что локальное охлаждение или нагревание в промежуточном мозгу, средоточии центральных терморецепторов, может изменить пищевое поведение, как это предсказывает гипотеза, но не исключены и другие толкования тех же самых данных.
Чрезмерное потребление пищи ведет к отложению жира в тканях, а когда пищи недостаточно, жировые отложения используются. Если предположить существование липорецепторов, то о таких отклонениях от идеального веса тела могли бы сигнализировать промежуточные продукты жирового обмена, которые появляются при отложении или использовании жира; так могли бы возникать сигналы чувства голода или сытости (рис. 9-4). В пользу липостатической гипотезы говорят не которые убедительные экспериментальные данные, в частности упомянутые выше данные о том, что после насильственного кормления животные едят меньше, чем контрольные, пока не израсходуются их жировые отложения.
Согласно этому толкованию, липостатический механизм голода действует главным образом при долговременной регуляции потребления пищи, а сокращения пустого желудка и глюкостатический механизм принимают участие прежде всего в кратковременной регуляции. Термостатический механизм, возможно, участвует и там, и там (см. рис. 9-4). При таком разнообразии физиологических механизмов, создающих ощущение голода, даже в самых сложных условиях это ощущение и пищевой драйв обеспечивают потребление пищи в должных количествах.

Рис. 9-4. Возникновение чувства голода. Связанные с этим рецепторы показаны под их адекватными стимулами.
Насыщение.Как и при питье, человек и животные прекращают поедать пищу задолго до того, как ее всасывание из пищеварительного тракта устранит тот дефицит энергии, который первоначально вызвал голод и потребление пищи. Все процессы, в результате которых животное перестает есть, имеют общее название насыщение. Как всякий знает по собственному опыту, ощущение того, что пищи съедено достаточно,-это нечто большее, чем только исчезновение голода; одним из других его проявлений (часть из которых связана с удовольствием) является отчетливое чувство переполнения, если пищи съедено слишком много. По мере того как после принятия пищи проходит время, ощущение насыщения постепенно ослабляется и в конце концов после более или менее долгого нейтрального периода вновь уступает место голоду. По аналогии с процессами, приводящими к утолению жажды, можно принять как предпосылку, что ощущение в начале насыщения преаб-сорбтивно - оно наступает до усвоения пищи, т.е. в результате процессов, связанных с самим актом еды, а происходящее позднее усвоение питательных веществ вызывает постабсорбтивное насыщение и препятствует немедленному возобновлению голода. Обратимся теперь к процессам, лежащим в основе этих двух видов насыщения.
Вероятно, преабсорбтивное насыщение создается многими факторами. Животные с фистулой пищевода, через которую проглоченная пища выходит, не попав в желудок, едят значительно дольше, чем до операции, и через более короткие интервалы. По-видимому, преабсорбтивно-му насыщению способствует стимуляция во время еды обонятельных, вкусовых и механорецепторов в слизистой носа, рта, глотки и пищевода, а возможно, также сам акт жевания (рис. 9-5), хотя имеющиеся в настоящее время данные указывают, что эти влияния на возникновение и сохранение чувства насыщения невелики. Еще одним фактором, по-видимому, является растяжение желудка пищей (рис. 9-5). Если желудок экспериментального животного наполнить через фистулу еще до того, как его накормят, то потом оно поедает меньше пищи. Степень компенсации связана не с питательной ценностью пищи, а с объемом первоначального содержимого желудка и с временем, когда оно туда введено. В крайних случаях потребление пищи через рот может быть полностью заторможено на недели, если в желудок непосредственно закладываются большие количества пищи незадолго до того, как животному полагается питаться. Следовательно, здесь безусловно играет роль растяжение желудка (и, возможно, прилегающей части кишечника). Наконец, хеморецепторы в желудке и верхних отделах тонкого кишечника (рис. 9-5),'очевидно, чувствительны к содержанию глюкозы и аминокислот в пище. Наличие соответствующих «глюкозных» и «аминокислотных» рецепторов в кишечной стенке показано электрофизиологически.

Рис.9-5. Возникновение чувства насыщения при приеме пищи. Жевательные движения могут способствовать преабсорбтивному насыщению через прямую центральную эфферентационную копию моторных программ жевания, через рецепторы, активи-руемые при жевании (например, мышечные веретена и сухожильные органы), или через те и другие.
Постабсорбтивное насыщение тоже может быть связано с указанными хеморецепторами, потому что они способны сигнализировать о концентрациях еще не использованных питательных веществ, оставшихся в пищеварительном тракте. К этому добавляются все энтероцептишм.к-сенсорные процессы, рассмотренные при обсуждении кратковременном и долговременной регуляции голода. Увеличение количества глюкозы и усиление теплопродукции по мере переработки пищи, а также изменения жирового обмена действуют на соответствующие центральные рецепторы (рис. 9-5); возникающие эффекты противоположны тем, которые вызывают голод (обозначены красным на рис. 9-4). В этом смысле голод и насыщение составляют две стороны одной монеты. Чувство голода побуждает к еде, а чувство насыщения (преабсорбтивного) заставляет прекратить еду. Однако количество съеденной пищи и продолжительность пауз между приемами пищи определяются также процессами, которые мы назвали «долговременной регуляцией приема пищи» и «постабсорбционным насыщением»,-процессами, которые, как мы теперь понимаем, в большей или меньшей степени накладываются друг на друга (ср. рис. 9-4 и 9-5).
Психологические факторы, участвующие в регуляции потребления пищи. В дополнение к перечисленным физиологическим факторам в регуляции пищевого поведения участвует ряд психологических факторов. Например, время приема пищи и количество съеденной пищи зависят не только от чувства голода, но также от установившихся привычек, количества предлагаемой пищи, ее вкуса и т. п. Животные, как и человек, регулируют количество поедаемого корма в зависимости от того, когда ожидается следующее кормление, и от того, сколько энергии, вероятно, будет до этого затрачено. Этот элемент планирования пищевого поведения, благодаря которому энергия поступает заранее, аналогичен «вторичному питью» (см. разд. 9-1), т.е. обычному порядку потребления воды.
Наше желание поесть определенную пищу, т.е. стремление повторить полученное удовольствие, называется аппетитом. Он может возникать из ощущения голода (т. е. пищевого драйва) или же появляться независимо (когда человек видит или ему предлагают что-нибудь особенно вкусное). Аппетит часто имеет соматаческую основу - например, стремление к солёной пище; когда организм потерял много соли; но он может также не зависеть от физических потребностей и отражать врожденные или приобретенные индивидуальные предпочтения. Такое приобретенное поведение, а также отказ от некоторых видов пищи объясняется наличием той или иной пищи и установившимися привычками, порой связанными с религиозными соображениями. С этой точки зрения «аппетитность» блюда - основные элементы которой составляют запах, вкус, консистенция, температура и то, как оно приготовлено и подано, - в большой мере зависит от нашей аффективной реакции на него. Примеров этому много, и их легко найти на местном, национальном и международном уровнях.
Почти любой человек, оказавшись перед соблазнительной пищей, иной раз съест ее больше, чем нужно. Механизмы кратковременной регуляции тут не справляются. После этого надо было бы сократить прием пищи, но в современном материально обеспеченном обществе не все ведут себя так. Причины несостоятельности долговременной регуляции, к сожалению, мало понятны. Программы предупреждения тучности, и борьбы с ней трудно разработать, и часто они не дают результатов; тучность и все связанные с ней опасности для здоровья во многих западных странах достигли, по-видимому, эпидемических размеров.
В заключение надо указать на связь между потреблением пищи и не-врозами и психозами. Усиленная еда или отказ от пищи часто служат эквивалентом удовольствия или протеста у психически больных, когда на самом деле беспокойство связано с другими видами мотивации. Наиболее известным примером является anorexia nervosa, форма отказа от пищи, весьма распространенная у девочек в период созревания; это нарушение психического развития может быть настолько серьезным, что приводит к смерти от голода.
Центральные механизмы голода и насыщения. Гипоталамус-структура, тесно связанная с регуляцией вегетативных функций (см. разд. 9.1), по-видимому, является главной центральной перерабатывающей и интегрирующей структурой также для голода и насыщения. Двустороннее разрушение. ткани в определенных вентромедиальных.областях гипоталамуса вызывает у экспериментального животного чрезвычайную тучность в результате переедания. В то же время разрушение более латеральных областей может привестик_отказу_от пищи и в конце концов к смерти от голода. С этими данными сопоставимы результаты локальной стимуляции гипоталамуса через вживленные электроды и опыты с золототиоглюкозой. Таким образом, в течение некоторого времени внимание исследователей было направлено почти исключительно на гипоталамус. В результате роль других структур мозга в регуляции потребления пищи известна очень мало. Безусловно будет упрощением заключить на основании только что упомянутых опытов, что вся центральная переработка информации локализована в двух «центрах», один из которых действует как «центр насыщения», а другой как «центр голода». Согласно этой гипотезе, разрушение центра насыщения должно привести к растормаживанию центра голода и отсюда к развитию волчьего аппетита; если же разрушен центр голода, то это должно вызывать постоянное чувство насыщения и отказ от всякой пищи. Однако положение явно намного сложнее. Например, упомянутые выше еда и питье «авансом» связаны с участием высших уровней головного мозга (лимбической системы, ассоциативной.коры). Не следует также упускать из виду, что еда и питье-это сложные двигательные акты, которые соответственно требуют широкого участия двигательной системы.
Дата добавления: 2015-10-05; просмотров: 1953;