Психофизика зрительного восприятия
Вразделе, посвященном общей субъективной сенсорной физиологии, время, место, качество и интенсивность были представлены как основные размерности при описании любого восприятия. Далее эти основные размерности будут рассмотрены в приложении к некоторым примерам. Рассматривается влияние пространственных и временных свойств зрительных стимулов на видение; обсуждаются такие качества зрительного восприятия, как яркость, цвет и глубина.
«Собственный свет сетчатки»(«Eigengrau»). Если испытуемый долгое время находится в совершенно темной комнате, а затем пытается описать свое зрительное восприятие, то он говорит, что видит не «черное» или «темное» поле, а скорее сероватое с более светлыми «туманными пятнами», светящимися точками и даже некоторыми упорядоченными структурами, изменяющимися в пространстве и времени. Разные испытуемые неизменно говорят о сероватом поле (иногда называемом собственным светом сетчатки), тогда как дополнительные зрительные восприятия сильно варьируют. Предполагается, что ощущение серого является субъективным коррелятом спонтанной активности нервных клеток сетчатки и центрального отдела зрительной системы.
Серая шкала.Представим себе, что перед человеком, который сидит в темной комнате и описывает свое восприятие «собственного света сетчатки», находится шахматная доска. Если ее внезапно осветить источником света, достаточно слабым, чтобы не ослепить человека, он немедленно увидит доску «правильно»: черные квадраты будут казаться ему темнее, чем серое поле, которое он видел перед этим, а белые квадраты светлее, хотя теперь на все части сетчатки света падает больше, чем

Рис. 4-11. Одновременный контраст (см. текст).
н предыдущий период полной темноты. Это простое наблюдение показывает, что восприятие черного, серого или белого зависит не только от количества света, падающего на каждый рецептор в единицу времени, но также от относительной освещенности соседних участков сетчатки. При условиях освещения, преобладающих при нормальном дневном свете, можно различить 30-40 разных градаций серого-от совершенно черного до ярчайшего белого.
Поскольку восприятие «внутреннего света» очевидным образом характеризует «состояние покоя» зрительной системы, Геринг (Е. Hering) более 100 лет назад предложил интерпретировать восприятия черного^ и белого (в примере с шахматной доской) в понятиях двух,противоположно действующих нейронных систем; эти «антагонистические» системы могли находиться в сетчатке или же в высших зрительных центрах. С этой точки зрения субъективное восприятие яркости в любом участке поля зрения определяется относительной степенью активации темповой (черной) системы и Светловой (белой) системы. Нейронные системы, постулированные Герингом, действительно были обнаружены как в слое ганглиозных клеток сетчатки, так и в высших зрительных центрах. Эти элементы упоминаются в разд. 4.3; это нейроны с «оп»-центром (светловая система) и нейроны с «оп»-центром (темновая система).
Одновременный контраст.На рис. 4-11 показан одновременный зрительный контраст. Серый участок на белом фоне кажется темнее, чем такой же серый участок на черном фоне. Иными словами, свойства фона определяют субъективно воспринимаемую яркость серых полей даже при том, что они одинаковые. Вдоль гранцы между светом и темнотой светлая часть кажется особенно яркой, а более темная часть кажется темнее, чем на расстоянии от границы. Этот эффект называется «усилением краевого контраста», а участки измененного восприятия известны под названием «полос Маха». Увидеть полосы Маха на рис. 4-11 можно лучше всего, фиксируя середину одного из'серых дисков с расстояния около 50 см. При фиксации левого диска его край кажется ярче центральной части, а темная зона сразу же за краем кажется темнее остального фона.

|
Рис. 4-12. Возникновение последовательных обра-
Явление одновременного контраста показывает, что субъективная реакция на стимуляцию определенной части сетчатки зависит от освещения окружающих частей. Ниже будет показано, что эту зависимость можно объяснить функциональной организацией рецептивных полей нейронов с «on-» и «оff»-центрами.
 Последовательный контраст между светом и темнотой; последовательные образы.Реакция на возбуждение определенного участка сетчатки зависит не только от одновременного возбуждения соседних участков (как показано выше), но также от пространственного распределения освещенности в непосредственно предшествующий отрезок времени. Изменение чувствительности на ограниченном участке сетчатки называется локальной адаптацией. Чтобы наблюдать ее на вашей собственной сетчатке, фиксируйте в течение 12 с одним глазом белую точку в центре геометрической фигуры справа на рис. 4-12, а затем фиксируйте точку в середине соседнего круга. Вы увидите теперь «последовательный образ». Участки исходной фигуры, которые были темными, кажутся в повйтельном образе ярче фона в белом поле, а те части, которые были светлыми, теперь кажутся темнее. Очевидно, те места сетчатки, на которые попадали темные части фиксируемой фигуры, стали более чувствительными, чем соседние, сильнее освещаемые участки.
Последовательный контраст между светом и темнотой; последовательные образы.Реакция на возбуждение определенного участка сетчатки зависит не только от одновременного возбуждения соседних участков (как показано выше), но также от пространственного распределения освещенности в непосредственно предшествующий отрезок времени. Изменение чувствительности на ограниченном участке сетчатки называется локальной адаптацией. Чтобы наблюдать ее на вашей собственной сетчатке, фиксируйте в течение 12 с одним глазом белую точку в центре геометрической фигуры справа на рис. 4-12, а затем фиксируйте точку в середине соседнего круга. Вы увидите теперь «последовательный образ». Участки исходной фигуры, которые были темными, кажутся в повйтельном образе ярче фона в белом поле, а те части, которые были светлыми, теперь кажутся темнее. Очевидно, те места сетчатки, на которые попадали темные части фиксируемой фигуры, стали более чувствительными, чем соседние, сильнее освещаемые участки.
Острота зрения.Изображение бинокулярно фиксируемого объекта
|
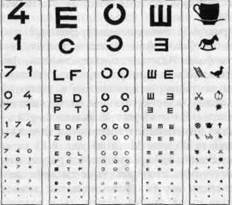
Рис. 4-13. Различные таблицы для проверки зрения с цифрами, буквами, кольцами Ландольта, знаками Снеллена и изображениями предметов (Leydhecker, 1975).
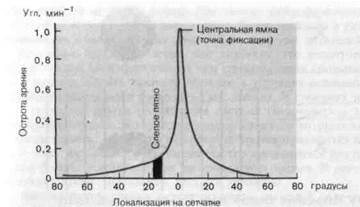
Рис. 4-14. Зависимость остроты зрения от положения тестирующего стимула на се [ чатке (фотопические условия).
проецируется, как упомянуто выше, на центральную ямку каждого гла-ia. При дневном свете острота зрения в этой части сетчатки наибольшая; по направлению к периферии сетчатки она постепенно снижается. В клинической практике острота зрения измеряется при помощи таблиц, подобных показанной на рис. 4-13; устанавливаются самые мелкие знаки, какие испытуемый видит отчетливо. Когда применяются кольца Ландольта, острота зрения выражается (как на рис. 4-14 и 4-15) it виде 1/а, где а-угол (в угловых минутах), соответствующий наименьшему различаемому разрыву в кольце. Другой мерой остроты зрения служит отношение Снеллена (Snellen): в числителе здесь стоит расстояние между наблюдателем и таблицей с буквами, обычно это 20 футов
| Рис. 4-15. Зависимость остроты фения от состояния адаптации сетчатки. Стрелкой показана область перехода от фотопиче-ских условий адаптации к ско-i опическим. |

Рис. 4-16. Нахождение слепого пятна правого глаза (см. текст).
(6,1 м). Знаменатель вычисляется после определения величины наименьших читаемых букв и представляет собой расстояние, на котором черные полоски, образующие эти буквы, составляют угол Г. Для людей с «нормальным» зрением эти расстояния равны; отсюда острота их зрения выражается отношением 20/20.
Остроту зрения можно измерять для разных частей сетчатки, если наблюдатель фиксирует одним глазом точку вне таблицы. На рис. 4-14 показано измеренное отношение между остротой зрения и положением на сетчатке. Следует также указать, что на остроту зрения влияет яркость рассматриваемого знака. На рис. 4-15 показана зависимость остроты зрения в фовеальной области (близ точки фиксации) от средней освещенности предъявляемой фигуры. Все знают из собственного опыта, что острота зрения зависит от условий освещения:
Вы не могли бы прочесть этот мелкий шрифт без достаточного освещения.
Правильный выбор освещения важен не только для чтения, но и на производстве и при домашней работе.
Слепое пятно.При внимательном изучении рис. 4-4 видно, что часть внутренней задней поверхности глаза не покрыта сетчаткой; это сосок, или место выхода зрительного нерва. Часть поля зрения (рис. 4-2), проецирующаяся на сосок,-это слепое пятно. В области слепого пятна не видно «ничего». Чтобы убедиться в существовании у вас слепого пятна, посмотрите на рис. 4-16 и фиксируйте одним правым глазом верхний крест с расстояния около 25 см. Вы заметите, что черный диск справа «исчез»; его изображение попадает на сосок вашего правого глаза.
Но нижняя часть рис. 4-16 показывает, что, хотя стимул на слепом пятне не виден, но область слепого пятна «заполнена», на нее экстраполируется изображение с окружающего участка. Когда вы фиксируете нижний крест на этом рисунке (одним правым глазом на расстоянии около 25 см), то изображение мыши исчезает в слепом пятне, но в вертикальной решетке не будет дырки. Как можно нейрофизиологически объяснить такое «перцептивное заполнение», будет показано в разд. 4.3.
Фотопическое и скотопическое зрение.Большинство читателей помнит из школьных курсов биологии, что колбочки-это рецепторы, которыми мы видим преимущественно при дневном свете (фотопическое), а палочки служат главным образом в условиях освещения, сооветствующих ясной_ночи (скотопическое зрение). В сумерках не преобладает ни та, ни другая рецепторная система (мезопическое зрение). Острота зрения наилучшая в фотопических условиях, в которых хорошо видны также и цвета; максимальная острота зрения приходится на центральную ямку. Временное разрешение быстро меняющихся фигур тоже лучше при фотопическом, чем при скотопическом зрении. В скотопиче-ских условиях время зрительных реакций удлинено.
При скотопическом зрении, осуществляемом системой палочек, преобладает функциональная цветовая слепота («ночью все кошки серы»), а острота зрения в области центральной ямки значительно хуже, чем при фотопическом зрении. Кроме того, положение наибольшей остроты зрения и максимальной чувствительности в скотопических условиях приходится не на середину центральной ямки, а на ее край. Это смещение точки максимальной чувствительности с фиксационной точки легко обнаружить, глядя ночью на звездное небо.
Если фиксировать взором очень бледную звезду, можно заметить, что она исчезает и появляется вновь, если избрать фиксируемую точку слегка в сторону от нее.
Процессы световой и темновой адаптации во времени.Если выйти ночью из ярко освещенной комнаты и пройти в сад, освещенный только звездами, то сначала вы практически не увидите ничего. Но постепенно чувствительность вашей зрительной системы приспособится к низкой интенсивности света в окружающей среде (темновая адаптация). По мере развития темновой адаптации острота зрения постепенно увеличивается и объекты в саду становятся видными хотя бы приблизительно.
Развитие темновой адаптации во времени можно выразить количественно, измеряя пороговую интенсивность стимулов, предъявляемых в разное время после перехода от света к темноте (рис. 4-17). Полученная таким образом кривая темновой адаптации показывает, что глаз не достигает максимальной чувствительности, пока не пробудет в темноте более 30 мин. Нормальная кривая темновой адаптации состоит из двух частей-первая считается колбочковои, вторая –палочковой.Колбочковый компонент можно измерить отдельно при помощи небольших пятен красного света, сфокусированных в фовеальной области. Хотя начальная адаптация (колбочковая) происходит быстрее, чем в палочках, конечная колбочковая чувствительность значительно ниже (т.е. пороги выше), чем палочковая (измеряемая белым или синим светом вне центральной ямки).
Процессом, противоположным темновой адаптации, является световая адаптация. Быстрое течение световой адаптации легко наблюдать,
 Рис. 4-17. Кривые темно вой адаптации. Прерывистая красная линия соответствует данным, полученным у полностью цветнослепого наблюдателя (палочковый монохромат); прерывистая черная линия получена при освещении красным светом центральной ямки, а черная сплошная — при освещении белым светом экстрафовеальной области у нормального испытуемого.
Рис. 4-17. Кривые темно вой адаптации. Прерывистая красная линия соответствует данным, полученным у полностью цветнослепого наблюдателя (палочковый монохромат); прерывистая черная линия получена при освещении красным светом центральной ямки, а черная сплошная — при освещении белым светом экстрафовеальной области у нормального испытуемого.
если, проведя много времени в темноте, войти затем в ярко освещенное помещение. При очень большой разнице в интенсивности вы будете на время ослеплены; затем за 15-60 с зрительная система приспосабливается к новой интенсивности света. Такого рода слепота связана с пониженной чувствительностью и нарушением восприятия формы. Подобное состояние может наступить, если, едучи ночью по шоссе, встретить машину с зажженными фарами.
Временные характеристики зрительного восприятия. Если смотреть на спицы колес быстро движущегося велосипеда, видна «прозрачная» серая поверхность. Смены света и темноты в движущихся изображениях спиц на сетчатке слишком часты, чтобы их заметить. Это обыденное наблюдение показывает, что существует верхний предел временного зрительного разрешения, называемый частотой слияния мельканий. Прерывистый световой стимул, мелькающий с частотой выше частоты слияния мельканий, неотличим от непрерывного света той же средней интенсивности. Частоту слияния мельканий легко измерить при помощи диска, разделенного на черные и белые секторы, как показано на рис. 4-18. Наблюдатель смотрит на часть такого диска, вращающегося все быстрее, пока не увидит его равномерно серым. Частота слияния мельканий возрастает с увеличением средней освещенности и размеров мелькающего светового стимула (рис. 4-18). Кривая на рис. 4-18 состоит из двух разных отрезков; часть ее, возрастающая более медленно, связана с палочковым зрением, а часть с более крутым наклоном-с колбочковым. Мелькающий свет воспринимается как таковой при частотах ниже 22 Гц в скотопических условиях адаптации и стимуляции. Более высокая частота слияния мельканий колбочковой системы говорит о том, что верецяя^щгнала^ колбочками происходит значительно быстрее, чем в палочках.
Обычно при измерении частоты слияния мельканий при каждом промере проверяется одна определенная часть поля зрения. Но если

Рис. 4-18. Зависимость частоты слияния мельканий от средней интенсивности светового стимула. Измерение при помощи синусоидально модулированного света; амплитуда модуляции - 70%.
проецировать последовательные световые стимулы попеременно на разные части сетчатки, то при этом вовлекаются временные и пространственные свойства переработки сигнала в сетчатке и в зрительной системе. Предположим, что в момент времени t0 световой стимул появляется в точке А поля зрения, а в момент t0 + tr он выключается; затем стимул вновь возникает (в момент времени t0 + tr + ∆t) в точке Б. В этих условиях наблюдатель воспринимает мнимое движение света от А к Б, неотличимqe от истинного движения, если ∆t < 120 мс. Попеременное освещение участков А и Б создает впечатление, будто световой стимул движется взад и вперед между А и Б. Описанное здесь явление мнимого движения для случая двух простых стимулов используется в кино. Проецируемая пленка состоит из последовательности неподвижных изображений разной конфигурации; они возникают друг за другом на экране с частотой от 18 до 24 кадров в 1 с. Такой частоты достаточно, чтобы вызвать в зрительной системе впечатление мнимого движения, при котором смена образов неотличима от истинного движения.
Цветовое зрение. Для человека с нормальным зрением предметы вокруг него обладают большим разнообразием цветовых качеств, или хроматическим рядом. Удобно рассматривать ступени серого как эквивалентный ряд, называемый также ахроматическим рядом. Он идет от ярчайшего белого до самого глубокого черного, создаваемого одновре- ., менным контрастом. Воспринимаемая окраска поверхностей объектов характеризуется тремя объективными величинами: цветовым тоном, насыщенностью и светлотой. Цвета располагаются в естественной замкнутой последовательности («цветовой круг» на рис. 4-19): красный, оранжевый, желтый, зеленый, синий, фиолетовый, пурпурный, красный.

Рис. 4-19. Положение цветов на цветовом круге. Цвета между (А) и (Б) не спектральные, а получены смешением красного и синего.


Рис. 4-20. Схема аддитивного (А) и субтрактивного (Б) смешения цветов.
Насыщенность цвета зависит от относительной величины хроматических и ахроматических компонентов. (Хроматичность, или окраска, определяется цветовым тоном и насыщенностью. Например, смесь спектрального красного с черным создает коричневый, а розовый создается смешением красного с белым.) Светлота указывает на относительное положение ахроматического компонента на шкале серого, между белым и черным.
Все цветовые тона в цветовом круге или соответствуют опреде-ненным цветам в спектре видимого солнечного света, или создаются аддитивным смешением двух спектральных цветов. Такое смешение происходит, когда свет разных длин волн падает на одну и ту же часть сетчатки (рис. 4-20). Для человека с нормальным цветовым зрением любой цвет, создаваемый освещенными предметами, может быть воспроизведен определенным аддитивным смешением трех надлежащим обра-юм выбранных монохроматических стимулов С1; С2 и С3. Такая ситуация полностью и однозначно описывается уравнением ощущения, или цветовым уравнением:
a[C1] + b[C2] + c[C3]~d[cx]. [4-2]
В этом выражении символ ~ означает «эквивалентность ощущения» или «подравнивание» цветов.
По международному соглашению, три определенные длины волны, основные цвета, обозначают С1 С2 и С3. Это спектральные цвета с длиной волны 700 нм (красный), 546 нм (зеленый) и 435 нм (синий); определенными их соотношениями "можно характеризовать любой цветовой оттенок.
Цвета можно также создавать процессом, противоположным аддитивному сочетанию. Такое субтрактивное (вычитательное) сочетание происходит, когда белый свет проходит "от источника через один (или более) цветовой фильтр (рис. 4-20). Когда художник смешивает краски (например, синюю и желтую, чтобы получить зеленую), он до некоторой степени использует субтрактивное смешение цветов, поскольку отдельные гранулы синей и желтой красок действуют как цветовые фильтры. Но при разглядывании красок на мозаиках или цветных изображениях известную роль может играть также аддитивное смешение, если две разные маленькие цветные точки попадают на одну группу колбочек. В «точечной манере» некоторых импрессионистов и в работах иуантилистекой школы систематически использовался этот эффект. Слияние цветов происходит только при достаточно большом расстоянии между картиной и наблюдателем.
Белое или черное как цвета.Для каждого цвета C1 в цветовом круге имеется второй цвет С2, который при аддитивном смешении с С1 дает белый цвет, т.е.
a [C] + b [C2] ~ W (белый). [4-3]
В этом цветовом уравнении смешения цветов коэффициенты а и b зависят от определения «белого». Цвета С1 и С2 во всех случах являются дополнительными. Белый свет может исходить от светящихся источников, но это невозможно для черного и разных оттенков серого-для их возникновения нужен механизм контраста. Термин цветовое тело применяется для представления воспринимаемых цветов в трехмерном пространстве. В простейшем цветовом теле его ось (черное/белое) «перпендикулярна» плоскости цветового круга, представляющего чистые цветовые тона.
Результаты опытов по сенсорной психологии с аддитивным смешением цветов [4-2] и [4-3] представлены геометрически двумерным «цветовым треугольником» (или диаграммой хроматичности, рис. 4-21). Если два цвета, входящие в треугольник, сочетаются аддитивно, то возникающий при этом цвет лежит на линии между этими двумя цветами. Цвет, дополнительный к любому другому, лежит на линии, проходящей через белую точку (Е на рис. 4-21). Для человека с нормальным цветовым зрением все цветовые тона излучений (выраженные уравнением [4-2]) удовлетворительно описываются не более чем тремя величинами. Это наблюдение породило термин «трихроматическое зрение». Значения коэффициентов a, b и с в уравнении [4-2], которые требуются для получения любого из цветов в цветовом треугольнике, практически одинаковы для преобладающей части населения; такие люди называются нормальными трихроматами. Для небольшого процента людей коэффициенты иные, и такие люди называются аномальными трихроматами. Около 1% людей -дихроматы; они воспринимают все цвета таким образом, что это можно описать уравнением с двумя членами:
a[C1] + b[c2]~d[Cx]. [4-4]
Диаграмма цветов, описывающая восприятие у этих людей, значительно менее дифференцирована, чем у трихроматов. Дихроматические отклонения от нормального цветового зрения, как и случаи аномальных трихроматов, обусловлены генетически. Самая обычная форма дихро-мазии-это «красно-зеленая слепота» (подробнее см. Ruch, Patton). Менее 0,01% людей полностью цветнослепые. Они различают только разные оттенки серого и называются монохроматами. Гены большинства полностью цветнослепых обусловливают существование у них только одной пигментной системы: их палочки и колбочки содержат только один пигмент-родопсин (см. разд. 4-3).
 Здесь следует сказать о двух главных теориях цветового зрения. Трихроматическая теория Юнга и Гельмгольца (R. Young, H. Helmholtz) постулирует существование трех типов колбочек, которые действуют как независимые системы приемников в фотопическом зрении; их сигналь! анализируются совместно нейронными системами восприятия яркости и цвета. Эту теорию подтверждают результаты по смешению цветов (уравнение [4-2]), открытие трех разных колбочковых пигментов(см. разд. 4.3) и различная спектральная чувствительность рецепторных потенциалов разных колбочек (см. рис. 4-26).
Здесь следует сказать о двух главных теориях цветового зрения. Трихроматическая теория Юнга и Гельмгольца (R. Young, H. Helmholtz) постулирует существование трех типов колбочек, которые действуют как независимые системы приемников в фотопическом зрении; их сигналь! анализируются совместно нейронными системами восприятия яркости и цвета. Эту теорию подтверждают результаты по смешению цветов (уравнение [4-2]), открытие трех разных колбочковых пигментов(см. разд. 4.3) и различная спектральная чувствительность рецепторных потенциалов разных колбочек (см. рис. 4-26).
Рис. 4-21. Цветовой треугольник, основанный на немецком стандарте DIN 5033. Ьелая зона окружает точку Е. Основание «цветового треугольника» образовано пурпурными тонами, которых нет в спектре и которые созданы аддитивным смешением спектральных цветов синего и красного.
Оппонентная теория цветов Э. Геринга (Е. Hering) постулирует существование антагонистических нейронных процессов для оппонентных цветов зеленый/красный и желтый/синий в дополнение к системе черный/белый, которая также организована антагонистически. Известная организация (см. разд. 4.3) рецептивных полей цветоспецифических нейронов сетчатки и латерального коленчатого тела, а также многие данные, полученные в опытах по сенсорной психологии, служат подтверждением этой теории. Таким образом, и трихроматическая теория цветового зрения и теория оппонентных цветов-Обе правильные на разных уровнях афферентной зрительной системы. На возможность синтеза этих двух главных теорий цветового зрения указывал еще в начале века в своей зональной теории фрейбургский физиолог Крис.
Бинокулярное зрение.Видимый объект представляется нам одним и тем же независимо от того, смотрим ли мы одним правым, одним левым или же обоими глазами. Объединение обоих монокулярных изображений объекта называется бинокулярным слиянием. По сравнению с монокулярным при бинокулярном зрении усилено впечатление пространственной глубины. Поскольку глаза расположены на голове не в одной точке, в изображениях окружающего мира, попадающих на обе сетчатки, имеются небольшие геометрические различия. Эти различия между изображениями, создаваемыми одним фиксируемым объектом на правой и левой сетчатках, тем сильнее, чем объект ближе. Различие (диспаратность) двух изображений служит предпосылкой восприятия глубины (стереоскопии). Чтобы достичь известной степени восприятия глубины при зрении только одним глазом, можно использовать различия в величине, степень перекрывания, параллакс, вызванный движением головы, но результат будет менее информативным, чем при полном бинокулярном зрении.
Разницу в изображениях видимого мира на обеих сетчатках можно продемонстрировать в следующем опыте: протяните правую руку во всю длину, поднимите большой палец и фиксируйте его сначала бино-кулярно, затем монокулярно попеременно правым и левым глазом. При смене глаз палец как бы перемещается по отношению к фону. Отсюда следует, что изображение окружающего мира на левой и правой сетчатке заметно различаются.
Бинокулярное слияние легко нарушается, если при фиксации объекта изменить положение одного глаза в глазнице, слегка нажав пальцем на веко. При этом объект проецируется на смещенную сетчатку «неправильно»: изображение «двоится», так как нарушено бинокулярное слияние.
Простое бинокулярное зрение (т.е. рассматривание одновременно только одного объекта) осуществляется лишь в том случае, когда изображения бинокулярно фиксируемого объекта падают на совершенно определенные части сетчатки в обоих глазах, на корреспондирующие участки сетчатки. При длительной бинокулярной фиксации данной точки (например, точки, отмеченной на рис. 4-22) можно определить эти корреспондирующие участки, передвигая тест-объект и находя те положения в пространстве, в которых он виден как один. Оказывается (как можно убедиться, начертив более подробные геометрические схемы), что все такие положения в пространстве падают на трехмерную изогнутую поверхность, включающую фиксационную точку. Эта поверхность называется гороптером, и, разумеется, для каждой фиксационной точки имеется свой гороптер. Для одной только горизонтальной плоскости ситуация проще-получаемая часть поверхности гороптера это «гороп-терный круг», показанный на рис. 4-22.
На заметном расстоянии от гороптера предметы «двоятся». Например, на рис. 4-22 при бинокулярной фиксации указанной точки обозначенный предмет лежит вне гороптера. Его изображения в обоих глазах явно падают на некорреспондирующие участки, а именно на противоположные стороны центральных ямок (углы а и Р на рис. 4-22).
Один из способов рассматривать проблему «глаза два, а мир один» состоит при таких ситуациях в том, чтобы представить себе «циклопический глаз», расположенный на продолжении гороптера посередине между глазами. Как показано на рис. 4-22, когда оба глаза, фиксирующие точку, отмеченную на гороптере, смотрят на объект, отдаленный от нее, то циклопический глаз увидел бы два мнимых предмета по обе стороны от реального. Когда же циклопический глаз видит только одно изображение, оно должно быть на гороптере, т.е. пары корреспондирующих участков в двух глазах по определению приходятся на одни и те же участки циклопического глаза.

Рис. 4-22. Схема бинокулярного зрения и построение циклопического глаза, видимый объект лежит здесь снаружи от поверхности гороптера. Его изображение проецируется справа от центральной ямки в левом глазу. При бинокулярном зрении создаются неперекрещивающиеся двойные изображения объекта; их положение можно определить, проецируя правую и левую сетчатки на воображаемую сетчатку циклопического глаза.
Существенное различие между изображениями двух объектов, лежа щих внутри и вне гороптера, легко продемонстрировать в «опыте с двумя пальцами». Держите перед собой правый указательный палец на уровне глаз на расстоянии около 20 см, а левый палец - за ним, на расстоянии около 50 см от глаз. Сначала фиксируйте ближний палец, тогда дальний будет двоиться. Затем закрывайте попеременно правый и левый глаз, и вы увидите, что двойные изображения не перекрещиваются. Это значит, что левый глаз видит левое изображение, а правый-правое. Дальний палец лежит снаружи от гороптера, создаваемого фиксацией ближнего пальца. Затем фиксируйте дальний палец. Теперь двоится ближний, и, попеременно закрывая то один, то другой глаз, вы увидите, что двойные изображения перекрещиваются: правый глаз видит левое изображение, и наоборот. Теперь ближний палец находится внутри гороптера, определяемого фиксацией дальнего пальца. Попробуйте зарисовать эту ситуацию и построить положения возникающих изображений по аналогии с рис. 4-22.
На основании описанных наблюдений можно было бы заключить, что все видимые предметы, изображения которых не попадают на корреспондирующие участки сетчатки, потому что они лежат внутри или снаружи от гороптера, должны вызывать двойные образы. Но если вы глядите вокруг себя в хорошо структурированном видимом мире, то мешающее двоение не возникает. По-видимому, существуют механизмы, которые препятствуют этому. Один из таких механизмов состоит в низкой остроте зрения на периферии сетчатки. Второй, как полагают, представляет собой бинокулярный тормозный механизм в центральных отделах зрительной системы, подавляющий восприятие того или другого из двух диспаратных изображений. Глядя на разные объекты центральными ямками левого и правого глаза, можно наблюдать этот бинокулярный тормозный механизм. Для этого надо сделать две трубки длиной около 30 см и диаметром 3-4 см. В одну из них смотрите одним глазом на марку, в другую (другим глазом)-на монету. С этими резко различными стимулами бинокулярного слияния не происходит-вы не увидите объединенную «марко-монету». Благодаря бинокулярному тормозному механизму возникает «бинокулярная конкуренция», попеременное восприятие то марки, то монеты. Отдельные части обоих стимулов могут быть видны одновременно, но только рядом, а не наложенные одна на другую. В этой бинокулярной конкуренции контуры, как правило, эффективнее равномерно затемненных поверхностей. Это можно объяснить известной из опытов более выраженной активацией нейронов афферентной зрительной системы при действии контрастных границ (подробнее см. разд. 4.3) по сравнению с активацией однородными поверхностями.
Стереоскопическое зрение. Несмотря на то что от любого достаточно крупного трехмерного объекта, видимого бинокулярно, на центральные ямки обоих газ падают несколько разные изображения из-за рассмотренной выше геометрической ситуации, обычно такой объект не «двоится» в нашем зрении, мы воспринимаем его как расположенное в пространстве целое.

Рис. 4-23. Стереоскопическое восприятие, основанное на горизонтальной диспа-ратности.
Расхождения в горизонтальной плоскости между изображениями на двух сетчатках (когда голова в нормальном положении), т.е. отклонения от точно корреспондирующих участков сетчаток1, называются горизонтальной диспаратностъю рецептивных полей. Она служит важным условием бинокулярного трехмерного зрения. Степень диспаратности определяет силу восприятия глубины. Поэтому -приведенное выше описание функционального значения корреспондирующих участков сетчаток следует видоизменить: только в случае, когда определенная горизонтальная диспаратность превышена, восприятие бинокулярно видимого объекта как единого целого уступает место двоящемуся образу. Иными словами, восприятие объекта как единого при стереоскопическом зрении требует функционального анализа не корреспондирующих точек в сетчатках обоих глаз, а связанных между собой изображений, падающих на участки в несколько угловых градусов. Горизонтальная диспаратность двух изображений предмета уменьшается по геометрическим причинам с увеличением расстояния от него до глаз. Поэтому на больших расстояниях точное восприятие глубины становится невозможным. Оптический дальномер увеличивает функционально расстояние между глазами и, кроме того, увеличивает размеры видимых предметов, улучшая тем самым восприятие глубины. Влияние горизонтальной диспаратности на восприятие глубины можно изучать при помощи рис. 4-23. Фиксируйте воображаемую точку за книгой так, чтобы оба больших круга бинокулярно слились. Тогда вы увидите три больших круга, средний из них-обоими глазами. При этом вам будет казаться, что маленький круг парит в пространстве перед плоскостью рисунка. Его изображения в глазах горизонтально диспаратны из-за асимметрии рисунка.
Восприятие формы. Если смотреть на хорошо структурированное окружающее пространство, то видны не изолированные «стимулы», .1 предметы в их соотношениях между собой. «Расчленение» механизмов фительного восприятия на простые процессы («сенсорные впечатления», см. с. 5), как это сделано в предыдущих разделах, оказалось по-иезным для сенсорной физиологии; это очевидный подход к количественному определению и пониманию функции периферических органов чувств. Гипотеза, связанная с таким подходом, а именно что восприятие сложной формы осуществляется элементарными физиологическими механизмами в сетчатке и афферентных путях зрительной системы, подтверждена прямыми нейрофизиологическими измерениями в некоторых системах (см. разд. 4.3).
 Рис. 4-24. Примеры иллюзий при восприятии формы, а-Расстояния АБ объективно одинаковые в изображениях двойной стрелки и двойной вилки, но они кажутся разными; б-две толстые вертикальные линии в действительности прямые и параллельные, в чем можно убедиться при помощи линейки; в-белый квадрат посередине создан субъективным «заполнением»; г-круги в центре одинаковой величины на обеих фигурах.
Рис. 4-24. Примеры иллюзий при восприятии формы, а-Расстояния АБ объективно одинаковые в изображениях двойной стрелки и двойной вилки, но они кажутся разными; б-две толстые вертикальные линии в действительности прямые и параллельные, в чем можно убедиться при помощи линейки; в-белый квадрат посередине создан субъективным «заполнением»; г-круги в центре одинаковой величины на обеих фигурах.
В то же время еще неизвестно, каким образом из обнаруженных до настоящего времени элементарных механизмов возникает восприятие вещественного объекта (Gestalt). Нет также объяснения «феноменологической инвариантности» воспринимаемых предметов. Этим термином обозначается тот хорошо известный факт, что чайник воспринимается как один и тот же чайник независимо от условий освещения, его положения и ориентации, а также от того, виден он одним или двумя глазами. Чайник воспринимается как целое, даже если видна только его часть.
В предметном восприятии так же, как в элементарных механизмах восприятия цвета и света или темноты, участвуют многочисленные процессы оценки контраста. При определенных условиях они создают иллюзии. Некоторые из таких иллюзий показаны на рис. 4-24. Воспринимаемая длина и кривизна линии определяются не только ее «объективными» характеристиками, но также длиной и углом линий рядом с ней (рис. 4-24, а, б). Контуры простых форм заполняются, и можно видеть, как край их продолжается через объективно пустое пространство (рис. 4-24, в). Воспринимаемые размеры фигуры зависят от размеров окружающих фигур (рис. 4-24, г). Систематические исследования в области гештальт-психологии, в особенности в первой четверти нашего века, выявили множество правил, управляющих зрительным восприятием сложных форм. Поскольку последние современные нейрофизиологические данные не дают достаточного объяснения этим правилам или вовсе не объясняют их, они не будут здесь приведены. Впрочем, в разд. 4.3 описаны некоторые нейрофизиологические факты, которые позволяют в первом приближении подойти к пониманию восприятия сложных зрительных форм.
В 4.6. Одновременный контраст между светлым и темным является
а) ошибкой суждения, которую легко избежать, сосредоточив внимание;
б) следствием нарушения диоптрического аппарата глаза;
в) механизмом, который приобретается обучением;
г) эффектом, вызываемым латеральным торможением в сетчатке и в центральной нервной системе.
В 4.7. Которое из следующих утверждений приложимо к остроте зрения у человека с нормальным зрением?
а) В условиях световой адаптации наибольшей остротой обладает центральная ямка.
б) Острота зрения при световой адаптации выше, чем при темновой.
в) Участок сетчатки с самым острым разрешением варьирует в зависимости от состояния адаптации.
г) Острота зрения в области слепого пятна равна 1,0'.
д) Остроту зрения можно измерить кольцами Ландольта.
е) Когда глаза ослеплены ярким светом, острота зрения пони
жена.
В 4.8. Какова острота зрения у человека, который правильно опознает кольцо Ландольта с промежутком а = 0,8'?
В 4.9. Какие механизмы нужны для нормального стереоскопического зрения (двумя глазами)?
а) Бинокулярное слияние.
б) Световая адаптация.
в) Темновая адаптация.
г) Бинокулярное торможение мешающих двойных изображений
(бинокулярная конкуренция).
д) Горизонтальная диспаратность.
е) Одинаковое цветовое восприятие в обоих глазах.
В 4.10. Корреспондирующими участками сетчатки мы называем
а) отношения между центром и периферией рецептивного поля;
б) участки в обеих сетчатках, которые при нормальном биноку
лярном зрении связаны между собой;
в) участки одной сетчатки, которые находятся на одинаковых
расстояниях от центральной ямки;
г) участки сетчатки, где имеются только колбочки.
В 4.11. Гороптер-это
а) область поля зрения, проецирующаяся на центральную ямку;
б) область бинокулярного поля зрения, проецирующаяся на обе
центральные ямки;
в) воображаемая поверхность в пространстве, каждая точка которой проецируется на геометрически соответствующие корреспондирующие участки на двух сетчатках;
г) область окружающей среды, лежащая вне бинокулярного по
ля зрения;
д) ни одно из приведенных утверждений не правильно.
В 4.12. Которое из следующих утверждений правильно?
а) Если три разных монохроматических стимула падают на один фоторецептор в глазу, происходит, аддитивное смешение цветов.
б) Если два разных монохроматических световых стимула падают на один фоторецептор в глазу, происходит субтрактивное смешение цветов.
в) Когда художник смешивает красную и желтую краски, чтобы получить оранжевую, он производит субстрактивное смешение цветов,
г) Когда художник смешивает желтую и синюю краски, чтобы получить зеленую, он производит аддитивное смешение цветов.
д) Набор всех цветов от световых источников, воспринимаемых человеком с нормальным зрением, описывается правильно и однозначно уравнением смешения цветов, включающим три спектральных цвета (первичных цвета).
Дата добавления: 2015-10-05; просмотров: 3351;