Изменение организации рецептивного поля при темновой адаптации.
Антагонистическую организацию РП можно объяснить наложением возбудительного и тормозного процессов в рецептивной единице ганглиозной клетки сетчатки. Рассуждение основано здесь на допущении, что процессы латерального возбуждения передаются преимущественно конвергенцией сигналов от нескольких биполярных клеток на одну ганглиозную- У тормозных процессов порог выше, чем у возбудительных, причем латеральные тормозные процессы передаются горизонтальными и амакриновыми клетками. Относительная сила возбуждения и торможения в рецептивной единице ганглиозной клетки сетчатки зависит от состояния адаптации, т.е. от среднего количества света, падающего на довольно большую область сетчатки.
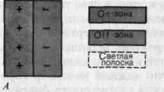
Рис. 4-30 иллюстрирует тот факт, что функциональный центр РП становится тем меньше, чем выше средняя освещенность предъявляемого паттерна, тогда как общая площадь РП остается постоянной. По мере развития темновой адаптации центр РП расширяется, и, когда она становится полной, антагонистическая организация рецептивных полей полностью исчезает. В этих условиях нейрон с «оп»-центром отвечает на освещение любой части своего рецептивного поля активацией при включении света, а нейрон с «оп»-центром отвечает по всему своему рецептивному полю активацией при выключении света.
 Рис. 4-30. Изменение величины центра РП нейрона с «оп»-центром в сетчатке млекопитающего при разных состояниях световой и темновой адаптации. При темновой адаптации периферия РП (серая зона) исчезает.
Рис. 4-30. Изменение величины центра РП нейрона с «оп»-центром в сетчатке млекопитающего при разных состояниях световой и темновой адаптации. При темновой адаптации периферия РП (серая зона) исчезает.
Плотность рецепторов и размеры РП как функция эксцентриситета на сетчатке.Эксцентриситет на сетчатке означает расстояние участка сетчатки от центральной ямки. Плотность рецепторов (их число на 1 мм2 поверхности сетчатки) меняется в зависимости от эксцентриситета. И колбочковой системе плотность рецепторов наибольшая в центральной ямке. Плотность же палочек наибольшая в парафовеальной области, а фовеа, центральная ямка, не содержит палочек. Плотность палочек также снижается по направлению к периферии сетчатки. На самом краю сетчатки рецепторный слой состоит почти полностью из палочек. Поскольку налочки не различают цветов, крайняя периферия функционально цветно-слепая.
Величина рецептивных полей растет от области центральной ямки к самому краю сетчатки. Данные, полученные на ганглиозных клетках светлоадаптированной сетчатки обезьяны, показывают, что центры рецептивных полей ганглиозных клеток в фовеальной области имеют и диаметре несколько угловых минут и увеличиваются по мере возрастания эксцентриситета. На периферии сетчатки центры РП ганглиозных клеток доходят до нескольких угловых градусов. Сравнение
истинной плотности рецепторов, полученной путем анатомических измерений, с размерами рецептивных полей позволяет думать, что конвергенция сигналов от рецепторов на ганглиозные клетки тоже растет с эксцентриситетом сетчатки. На основании тесной корреляции между диаметрами центров РП и остротой зрения в разных частях сетчатки можно заключить, что, как и следовало ожидать, острота зрения зависит не только от плотности рецепторов, но и от свойств1РП, Это заключение подтверждается тем что острота зрения изменяется при изменении средней освещенности; в этом случае диаметр центров рецептивных полей очень тесно коррелирует с остротой зрения, между тем как плотность рецепторов, разумеется, на зависит от состояния адаптации.
Ответы ганглиозных клеток на цветные стимулы. Количественная оценка поведенческих реакций говорит о том, что цветовое зрение у таких животных, как макак-резус, и у человека сходно. Однако функциональная организация рецептивных полей ганглиозных клеток сетчатки у этих животных сложнее, чем описанное выше деление на клетки с «on»- и «оп»-центрами. В светлоадаптированной сетчатке, а также в латеральном коленчатом теле и зрительной коре у них можно найти нервные клетки, дающие частично цвето-специфические ответы. Несколько упрощая, можно различить три класса таких клеток (рис. 4-31).
Ганглиозные клетки свето-темновой системы реагируют качественно одинаково (свойства «on»- и «он»-центров) при разных длинах волны монохроматического светового стимула в пределах видимого спектра (~ 400-700 нм). Спектральная чувствительность в центре и на периферии РП ганглиозных клеток этого класса одинакова.
Ганглиозные клетки красно-зеленой системы обладают частично цве-то-специфической антагонистической организацией РП. Монохроматические световые стимулы из красной области спектра вызывают активацию в центре РП и торможение на периферии, а монохроматические стимулы из зеленой области спектра вызывают ответы противоположного характера.
Ганглиозные клетки желто-синей системы при монохроматической стимуляции центра РП активируются желтым светом и тормозятся синим; наоборот, на периферии желтый свет тормозит, а синий возбуждает.
Таким образом, нейроны сетчатки и коленчатого тела осуществляют преобразование сигналов, имеющих отношение к цветовому зрению. Через биполярные, горизонтальные и амакриновые клетки сигналы от трех разных типов колбочек направляются таким образом, что создают в слое ганглиозных клеток одну нейронную систему для «ахроматического зрения» и две цвето-специфические антагонистические нейронные системы. Эти последние составляют четырехцветную оппонентную систему с оппонентными парами цветов желто-синей и красно-зеленой.
Проекции сетчатки на ЦНС. Зрительные нервы обоих глаз соединяются у основания черепа и образуют зрительную хиазму (перекрест) (рис. 4-32). В хиазме человека около полумиллиона волокон зрительного нерва переходят на противоположную сторону; вторая половина остается ипсилатеральной и вместе с перекрещенными аксонами второ-го зрительного нерва образует зрительный тракт. Следовательно, на каждой стороне около миллиона аксонов ганглиозных клеток сетчатки проходят в зрительном тракте к первым центральным переключа-юльным станциям зрительного пути к латеральному коленчатому телу,верхнему бугорку и претекталъной области ствола мозга. Перекрест нервных волокон в хиазме следует строгому правилу: аксоны ганглиозных клеток в височной части левой сетчатки и назальной
 Рис. 4-31. Схема про-странственной организации трех рецептивных полей в слое ганглиозных клеток сетчатки и в латеральном коленчатом теле млекопитающего с цветовым зрением.
Рис. 4-31. Схема про-странственной организации трех рецептивных полей в слое ганглиозных клеток сетчатки и в латеральном коленчатом теле млекопитающего с цветовым зрением.
 А. Нервная клетка в системе свет-темнота. Б. Нервная клетка в системе красный-зеленый. В. Нервная клетка в системе желтый-синий. И цветоспецифических рецептивных полях (Б и В) центр и периферия организованы антагонистически.
А. Нервная клетка в системе свет-темнота. Б. Нервная клетка в системе красный-зеленый. В. Нервная клетка в системе желтый-синий. И цветоспецифических рецептивных полях (Б и В) центр и периферия организованы антагонистически.

части правой сетчатки входят в левый зрительный тракт, а аксоны от ганглиозных клеток назальной части левого глаза и височной части правого глаза образуют правый зрительный тракт (рис. 4-32). Аксоны большей части нервных клеток латерального коленчатого тела проходят по зрительной радиации
 Рис. 4-32. Схема зрительных путей в головном мозгу человека, справа показаны также некоторые эфферентные связи между зрительной корой и подкорковыми структурами.
Рис. 4-32. Схема зрительных путей в головном мозгу человека, справа показаны также некоторые эфферентные связи между зрительной корой и подкорковыми структурами.
Зрительные зоны в левом и правом полушариях соединены между собой аксонами, идущими через мозолистое тело. Разделение на поля 17, 18 и 19 сильно упрощено.
к первичной зрительной коре (поле 17) в затылочной доле большого мозга. Эта область коры связана со вторичными и третичными зрительными центрами, а также высшими зрительными ассоциативными областями, тоже лежащими в затылочной области коры больших полушарий или в теменной.коре_. Из этих зрительных зон идут пути к корковым „станциям переключения афферентной зрительной системы,"показанным на"рис"" 4-32, к участкам ствола мозга, контролирующим движения глаз (см. разд. 4.4), и к ассоциативным зонам в теменной и височной долях, функция которых связана с речью. Зрительные центры левого и правого полушарий сообщаются через мозолистое тело (рис. 4-32).
Центральный зрительный путь характеризуется топографической организацией. Это значит, что подобно тому, как пространственные отношения в определенной географической области сохраняются на ее карте, так и
пространственное распределение возбуждения в слое ганглиозных клеток сетчатки «картируется» пространственным распределением возбуждения нейронов в латеральном коленчатом теле, верхнем бугорке и зрительной коре. Но детали такого картирования несколько отличны от географического стандарта. Карта чертится в определенном масштабе (например, уменьшение 1:100 000), так что каждый километр горизонтального расстояния естественной местности соответствует фиксированному расстоянию на бумаге, тогда как топографическая проекция сетчатки нелинейна. Маленькая область центральной ямки проецируется на гораздо большее число центральных нейронов, чем область такой же величины на периферии сетчатки. Эта нелинейная проекция отражает гораздо большее функциональное значение центральной ямки по сравнению с периферией, а также снижение остроты зрения по мере удаления от ямки (см. разд. 4.2).
Нейронная переработка сигналов в латеральном коленчатом теле. Нервные клетки латерального коленчатого тела, первой станции переключения на пути между сетчаткой и корой большого мозга, обычно обладают простыми концентрическими РП (судя по измерениям при стимуляции сетчатки светом), подобно РП ганлиозных клеток сетчатки . Бинокулярное взаимодействие между сигналами от левого и правого глазa выявляется здесь только как слабое реципрокное торможение. В трех из шести слоев нервных клеток коленчатого тела «доминирует» один глаз, но в остальных трех слоях-второй. Таким образом, в каждой из двух трехслойных групп происходит переработка сигналов преимущественно одного глаза. Истинная бинокулярная интеграция зрительной информации от обоих глаз происходит впервые в зрительной коре. Функция латерального коленчатого тела до сих пор далеко не ясна, несмотря на
обширные исследования. Имеются данные о том, что процессы латерального торможения, столь важные при различении контраста (см. разд. 4.2), особенно выражены в некоторых клетках коленчатого тела. И, как было указано выше, информация о цвете представлена здесь по меньшей мере в трех разных классах нервных клеток (светло-темной, желто-синей и красно-зеленой системах). Возбудимость нервных клеток " латеральном коленчатом теле сильно меняется с переходом от бодорствания ко сну. В состоянии глубокого сна передача сигнала в коленчатом теле, вероятно, резко снижена.
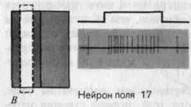
Нейроны зрительной коры. В нервных клетках первичной, вторичной ,третичной зрительных зон затылочной коры появляются такие особенности организации РП, которые неизвестны на нижних уровнях. Строение, типичное для нейронов сетчатки и коленчатого тела с концентрическими возбудительными и тормозными зонами, обнаружено лишь для некоторых нейронов первичной зрительной коры. В рецептивных Полях остальных корковых зрительных нейронов «on»- и «оff»-зоны расположены параллельно друг другу (рис. 4-33). Диффузное освещение всего РП обычно мало меняет спонтанную активность этих клеток. Но если на РП проецируется «полоска» света в правильных ориентации и положении, то наступает сильная активация. Так происходит, например, когда полоска ориентирована, как на рис. 4-33, В. РПс такими параллельными «on»- и «оff»-зонами называются «простыми», потому что их функциональная организация легко устанавливается путем проецирования маленьких пятнышек света на разные части РП; иначе говоря, ответ на полоски можно понять на основании ответов на пятнышки.
Другие же корковые зрительные нейроны обладают «сложными» РП.Для активации таких нейронов нужно проецировать на РПсветло-темные контуры со специфической пространственной ориентацией и размерами, перерывами линии, углами и т. п. Все участки РП,раздражение которых «правильным» стимулом вызывает активацию нейрона, называют возбудительным рецептивным полем (ВРП) Обычно ВРПокружено областью, в которой светло-темные стимулы вызывают только торможение. Эта область называется тормозным рецептивным полем (ТРП). Нейроны со сложными РПобычно не отвечают на проецируемые на их РПдиффузные неструктурированные световые стимулы. На рис. 4-34 показаны ответы двух нейронов зрительной коры со сложными РП.
|
|


Рис. 4-33. Организация «простого» рецептивного поля нервной клетки в первичной зрительной коре. (Упрощенный рисунок из Hubel, Wiesel, J. Physiol. (Lond.), 160,106-154, 1962.)
Многие корковые нейроны со сложными РПактивируются гораздо сильнее не неподвижными, а движущимися стимулами. Кроме того, некоторые из этих нейронов, чувствительных к движению, требуют определенного направления движения стимула. Например, стимул с оптимальной пространственной структурой окажет возбуждающий эффект, только если он движется по ВРПслева направо, а движение в обратном направлении не будет активировать клетку. Чувствительность корковых зрительных нейронов к движению и направлению, несомненно, отражает приспособление к тому, что изображение неподвижного окружения всегда смещается по сетчатке из-за непрерывного движения глаз и тела (см. разд. 4.4). «Мозговой образ» неподвижного зримого мира должен быть создан во время коротких периодов фиксации из картин на сетчатке, которые меняются при каждом движении глаза.

Рис. 4-34. Схема ответов двух нейронов в зрительной коре (поля 18 и 19) со сложными рецептивными полями. Стимул обозначен серой штриховкой, стрелка показывает направление движения. (Упрощенный рисунок из Hubel, Wiesl, J. Neurophysiol., 28, 229-289, 1965.)
Нейроны зрительной коры со сложными РП,как правило, обладают своими РПдля каждого глаза. Поэтому эти клетки могут быть возбуждены монокулярно, т.е. раздражением одного глаза. Бинокулярная стимуляция одинаковыми стимулами приводит к более сильной актива  ции (бинокулярная суммация). РП бинокулярно активируемой корковой нервной клетки, приходящиеся на участки левой и правой сетчатки, обладают только приблизительным геометрическим соответствием (см. разд. 4.2). Отклонение этих участков от точного соответствия областей сетчатки варьирует у разных нейронов. Эта диспаратность такова, что стимул, наносимый бинокулярно, вызывает оптимальное возбуждение, когда он лежит несколько снаружи от гороптера. Такой нейронный ответ рассматривается как нейрофизиологический коррелят бинокулярного восприятия глубины, о чем более подробно речь шла выше. Нейронная основа восприятия формы.Корковые зрительные нейроны со сложными РП отвечают только на узко специфические зрительные стимулы. Эти оптимально активирующие свойства различны для разных классов нейронов. В общей сложности в первичной, вторичной и третичной зрительной коре обнаружено не менее 15 классов нейронов; в каждом случае они распределены по всей корковой проекционной зоне поля зрения. Путем отведения активности по очереди от множества разных нервных клеток в зрительной коре можно реконструировать пространственную картину возбуждения, которая существует в данный момент в корковых нейронах одного класса под воздействием определенного зрительного стимула. На рис. 4-35 дана попытка такой реконструкции в упрощенном виде. Пространственное распределение возбуждения в нейронах разных классов отражает разные свойства стимула. Поскольку проекция сетчатки на зрительной коре нелинейна, это распределение во многих тысячах нейронов данного класса не такое простое, как показано на рис. 4-35. Данная схема имеет целью лишь показать одновременные многочисленные представительства одного изображения, которые проявляются в особом распределении возбуждения в нейронах зрительной коры разных классов. Рисунок помогает также дать читателю представление о сложности переработки сигналов, с которой приходится иметь дело в нейрофизиологических исследованиях отдельных нервных клеток зрительной коры млекопитающих. Нейронные механизмы, воссоздающие форму из множественных предъявлений зрительной информации в виде разных конфигураций возбуждения в зрительной коре, пока еще совершенно неясны. Но из наблюдений над больными, у которых нарушено зрительное восприятие формы и предметов (зрительная агнозия), известно, что для этих функций необходимы не только зрительные проекционные зоны затылочной коры больших полушарий, но также высшие ассоциативные зоны теменной коры.
ции (бинокулярная суммация). РП бинокулярно активируемой корковой нервной клетки, приходящиеся на участки левой и правой сетчатки, обладают только приблизительным геометрическим соответствием (см. разд. 4.2). Отклонение этих участков от точного соответствия областей сетчатки варьирует у разных нейронов. Эта диспаратность такова, что стимул, наносимый бинокулярно, вызывает оптимальное возбуждение, когда он лежит несколько снаружи от гороптера. Такой нейронный ответ рассматривается как нейрофизиологический коррелят бинокулярного восприятия глубины, о чем более подробно речь шла выше. Нейронная основа восприятия формы.Корковые зрительные нейроны со сложными РП отвечают только на узко специфические зрительные стимулы. Эти оптимально активирующие свойства различны для разных классов нейронов. В общей сложности в первичной, вторичной и третичной зрительной коре обнаружено не менее 15 классов нейронов; в каждом случае они распределены по всей корковой проекционной зоне поля зрения. Путем отведения активности по очереди от множества разных нервных клеток в зрительной коре можно реконструировать пространственную картину возбуждения, которая существует в данный момент в корковых нейронах одного класса под воздействием определенного зрительного стимула. На рис. 4-35 дана попытка такой реконструкции в упрощенном виде. Пространственное распределение возбуждения в нейронах разных классов отражает разные свойства стимула. Поскольку проекция сетчатки на зрительной коре нелинейна, это распределение во многих тысячах нейронов данного класса не такое простое, как показано на рис. 4-35. Данная схема имеет целью лишь показать одновременные многочисленные представительства одного изображения, которые проявляются в особом распределении возбуждения в нейронах зрительной коры разных классов. Рисунок помогает также дать читателю представление о сложности переработки сигналов, с которой приходится иметь дело в нейрофизиологических исследованиях отдельных нервных клеток зрительной коры млекопитающих. Нейронные механизмы, воссоздающие форму из множественных предъявлений зрительной информации в виде разных конфигураций возбуждения в зрительной коре, пока еще совершенно неясны. Но из наблюдений над больными, у которых нарушено зрительное восприятие формы и предметов (зрительная агнозия), известно, что для этих функций необходимы не только зрительные проекционные зоны затылочной коры больших полушарий, но также высшие ассоциативные зоны теменной коры.
В 4.13. Которое из следующих утверждений приложимо к рецепторным потенциалам отдельных колбочек в сетчатке позвоночных?
а) При освещении возникает деполяризационный рецепторный потенциал
б) При освещении возникает гиперполяризационный рецепторный потенциал.

Рис. 4-35. Схема процессов возбуждения, вызываемых светящейся буквой К в разных нейронных слоях сетчатки и центральной нервной системы.
а. Изображение буквы на сетчатке и пространственное распределение возбуждения в ре-цепторном слое сетчатки, б, в. Распределение возбуждения на выходе сетчатки в слое ганглиозных клеток. Возбуждение обозначено красными полосами, б. Нейроны с «оп»-центром. в. Нейроны с «оff»-центром. г. Распределение возбуждения в нейронном слое латерального коленчатого тела и некоторых нейронах зрительной коры. Контуры светящейся буквы вызывают возбуждение в нервных клетках, д-и. Распределение возбуждения в разных слоях нервных клеток различных классов в первичной, вторичной и третичной зрительной коре головного мозга. Здесь нервные клетки возбуждаются только контурами определенной ориентации, углами или разрывами линий. На рисунке нейробиологические отношения сильно упрощены; пространственное распространение возбуждения в разных слоях не связано линейно с размерами стимула.
в) При освещении мембранный потенциал рецептора не меняет
ся.
г) Рецепторный потенциал совершенно не связан с движением
ионов через мембрану.
д) Имеется линейная зависимость между амплитудой рецептор-
ного потенциала и интенсивностью стимула.
е) Имеется приблизительная логарифмическая зависимость между амплитудой рецепторного потенциала и интенсивностью стимула.
4.14. Которое из следующих утверждений приложимо к родопсину (Р)?
а) Р состоит из опсина и ретиналя.
б) Р состоит из гамма-глобулина и витамина А.
в) Р идентичен зрительному пигменту в колбочках.
г) Концентрация Р в палочках растет при темновой адаптации,
д) Концентрация Р в палочках снижается при темновой адаптации,
е) Раствор Р красный, но обесцвечивается после освещения.
В 4.15. Закончите правильно следующие фразы. Нейроны сетчатки с «оn»-центром (ганглиозные клетки) отвечают на освещение центра РП (активацией/торможением), тогда как затемнение центра РП ведет к (активации/торможению). Освещение периферии РП нейронов с «оn»-центром вызывает (активацию/торможение), а затемнение вызывает (активацию/торможение). Следовательно, центр и периферия РП организованы. При увеличении средней интенсивности света, падающего на сетчатку, диаметр центра РП становится (больше/меньше).
В 4.16. Которые из следующих утверждений правильны? Рецептивные поля ганглиозных клеток в сетчатке млекопитающих
а) в фовеальной области меньше, чем на периферии;
б) в среднем одинаковых размеров по всей сетчатке;
в) могут быть подразделены на функционально разные центр
и периферию в условиях световой адаптации;
г) могут быть обнаружены только у нейронов центральной
ямки;
д) иногда организованы в отношении спектральной чувствительности антагонистически;
е) не крупнее рецептивных полей одиночного фоторецептора.
В 4.17. Зная организацию рецептивных полей нервных клеток зрительной коры, можно понять, почему следующие свойства зрительных стимулов особенно важны для восприятия формы,
а) Средняя освещенность.
б) Контуры.
b) Разрыв контура.
г) Цвета.
д) Тот факт, что на сетчатке создается перевернутое изображение видимых предметов.
Дата добавления: 2015-10-05; просмотров: 1617;