МЕТАБОЛИЗМ МИОКАРДА В НОРМАЛЬНЫХ УСЛОВИЯХ И ПРИ ИШЕМИИ
В среднем сердце взрослого человека сокращается около 100.000 раз в сутки и перекачивает от 7 до 8 литров в минуту при систолическом артериальном давлении 120 мм рт. ст. Для выполнения такой огромной работы требуется хорошо сбалансированный энергетический обмен. Митохондриальный аппарат занимает в кардиомиоцитах до 40% от всего объема клетки, что значительно больше, чем в большинстве других клеток организма. Кроме того, кристы в митохондриях кардиомиоцитов, в которых локализуется дыхательная цепь - конечное звено окислительного фосфорилирования и образования аденозинтрифосфата (АТФ) - упакованы более плотно, чем в других клетках. В качестве энергетического субстрата клетки миокарда способны использовать глюкозу, жирные кислоты, кетоновые тела и аминокислоты, последовательно переходя от одного субстрата к другому в зависимости от энергетических потребностей, характера питания, доступности тех или иных субстратов и состояния миокарда. В нормальных условиях главным энергетическим субстратом являются жирные кислоты. При повышенной физической нагрузке в качестве энергетического субстрата может выступать молочная кислота, а при голодании – кетоновые тела. После приема пищи, богатой углеводами, под действием инсулина происходит увеличение поступления глюкозы в клетки миокарда, и она становится основным энергетическим субстратом, в то время как после приема жирной пищи до 100% всей энергии в кардиомиоцитах образуется в результате b-окисления жирных кислот.
Жирные кислоты поступают к клеткам миокарда в связанном виде с альбумином или в составе триглицеридов. Они включаются в реакции окисления в митохондриях, куда доставляются в виде КоА-эфиров с помощью транспортной карнитиновой системы, включающей ферменты карнитин-пальмитоилтрансферазы (КПТ-I и КПТ-II). Жирные кислоты с короткими цепями могут поступать в митохондрии и минуя систему карнитина. Параллельно с жирными кислотами в энергетическом обмене участвует и глюкоза, которая превращается в процессе гликолиза в пируват, а затем в ацетил-КоА. Ацетил-КоА включается в цикл трикарбоновых кислот (ЦТК; цикл Кребса) и используется для выработки кардиомиоцитами АТФ (рис. 1).
В условиях ишемии миокард переходит преимущественно на питание глюкозой, что связано с несколькими факторами. Так, при дефиците поступления субстратов и кислорода к клеткам, они начинают использовать для своего энергообеспечения гликоген, который содержится в ограниченном количестве в кардиомиоцитах. Кроме того, несмотря на высокую энергетическую ценность жирных кислот, для получения одинакового количества энергии при окислении глюкозы требуется кислорода меньше, чем при окислении жирных кислот. Это связано с тем, что жирные кислоты, в отличие от глюкозы, имеют в своей химической
 |
структуре очень мало атомов кислорода.
Так, при одинаковом потреблении кислорода в условиях окисления глюкозы образуется на 14% энергии АТФ больше, чем при окислении жирных кислот. Поскольку при ишемии наиболее важным патогенетическим фактором является дефицит кислорода, а не субстратов, такой «кислород-сберегающий» метаболизм является спасительным и сохраняет энергетический обмен на максимально возможном в данных условиях уровне. При дефиците кислорода в сердце происходит высвобождение катехоламинов из нервных окончаний, что приводит к активации фосфорилазы и фосфофруктокиназы и усилению анаэробного гликолиза. Образующаяся в процессе гликолиза энергия составляет всего 7% от необходимого количества и не позволяет длительно сохранять жизнеспособность миокарда. Запасы АТФ при ишемии начинают уменьшаться уже через несколько сокращений сердца; через 15 минут содержание АТФ составляет лишь около 35% от нормы, а через 40 минут – около 10%. Уровень креатинфосфата уменьшается гораздо быстрее и его количество резко снижается уже через несколько секунд.
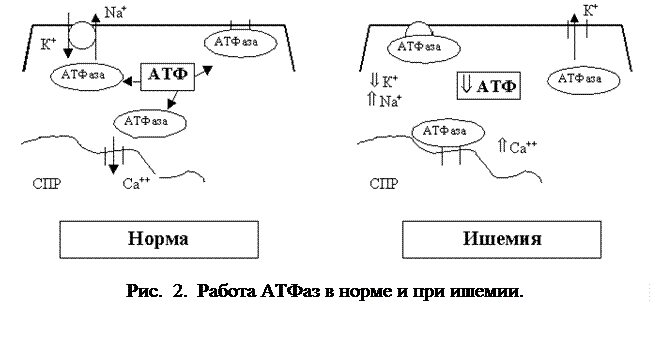
Расход энергии в кардиомиоцитах распределяется приблизительно в следующей пропорции: 80% - всей энергии АТФ идет на обеспечение сокращения сердца, 15% - на транспорт ионов через мембраны клеток и около 5% на синтетические процессы в клетках. Таким образом, основные пути утилизации энергии в кардиомиоците связаны с работой различных АТФаз, в частности: Са2+-АТФазы саркоплазматического ретикулума, Са2+-активируемой АТФазы миофибриллярного комплекса, Na+,K+- АТФазы, АТФазы К+ каналов и других. В последнее время большое значение в регуляции ионного транспорта при ишемии придается АТФ-чувствительным калиевым каналам, которые обеспечивают один из механизмов защиты кардиомиоцитов в условиях дефицита энергии. Особенностью этих каналов является то, что при нормальном энергетическом обмене клеток они не функционируют. В условиях дефицита энергии, например, при ишемии, напротив, происходит открытие этих каналов и калий начинает покидать клетку (рис. 2). При достаточном количестве энергии Na+, K+-АТФаза и Са2+-АТФаза обеспечивают транспорт ионов; АТФ-чувствительные К+- каналы не работают. При ишемии происходит открытие АТФ-чувствительных К+- каналов.
При этом сохраняется ионный гомеостаз и нормальные показатели осмолярности в клетке. При дефиците энергии нарушается работа Na+,K+-АТФ-азы, поэтому ионы натрия задерживаются в клетке и их внутриклеточная концентрация резко возрастает, что неизбежно приводит к чрезмерному возбуждению клеток, поскольку снижается (делается менее отрицательным) клеточный потенциал покоя. Открытие АТФ-зависимых К+ каналов приводит к выходу ионов калия из клетки, нормализации потенциала покоя и уменьшению возбудимости клеток.
Дата добавления: 2015-09-18; просмотров: 2503;