Поліаденілування мРНК і термінація транскрипції
Сигнал термінації транскрипції та поліаденілування (так званий polyA-сигнал) складається з двох елементів послідовності: консенсус AAUAAA і розташована в ~20 нуклеотидах нижче від нього U або G/U збагачена послідовність (рис. 8). Перший елемент упізнається гетеротетрамерним білком CPSF (Cleavage Polyadenylation Factor). Три із чотирьох субодиниць фактора є компонентами базального фактора транскрипції TFIID – вони переносяться на CTD після ініціації, і далі полімераза несе їх на собі, доки вони не зустрінуть сигнал. Другий елемент polyA-сигналу впізнається фактором розрізання РНК СstF (Cleavage stimulation Factor). Взаємодія обох факторів з РНК є кооперативною – вони підсилюють зв’язування одне одного.
Після первинного впізнання polyA-сигналу, поки РНК-полімераза продовжує синтез РНК за сигналом (до 1000 нуклеотидів), до мультибілкового комплексу, що збирається на polyA-сигналі, долучаються polyA-полімераза (РАР), ще два фактори розрізання (Сleavage Factors) CF 1 і 2 та, можливо, інші білки. Збирання комплексу стимулюється зв’язаним з кепом СВС, а також сплайсосомою на останньому інтроні – сплайсинг останнього інтрона та розрізання / поліаденілування РНК здійснюються одночасно і стимулюють одне одного. Зокрема, білок U2AF, який знаходиться на останньому інтроні, взаємодіє з РАР. У межах другого елемента послідовності polyA-сигналу знаходиться консервативний динуклеотид СА, в якому й відбувається розрізання (сleavage) РНК. Від якої конкретно активності залежить це розрізання, залишається невідомим. До 3′-кінця, що виник внаслідок розрізу, РАР (за стимулюючої дії CPSF) приєднує один за одним 100 – 200 аденінових нуклеотидів. З polyA-хвостом, що зростає, відразу зв’язується специфічний білок РАВР (PolyA Binding Protein), який підвищує процесивність РАР (рис. 8).
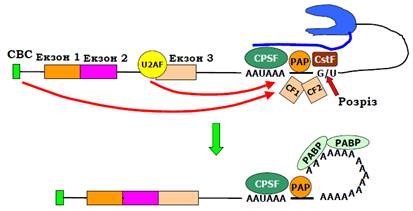
Рис. 8. Впізнання polyA-сигналу та поліаденілування мРНК.
Розрізання / поліаденілування РНК є тригером термінації транскрипції (рис. 9). Напевно, упізнання polyA-сигналу та розрізання / поліаденілування факторами, зв’язаними з РНК-полімеразою, індукує конформаційні зміни в полімеразному комплексі, які приводять до зниження спорідненості полімерази до ДНК та / або транскрипту. Крім того, процес розрізання та поліаденілування синхронізован із так званою ко-транскрипційною деградацією РНК-нуклеазами нижче від polyA-сигналу. Механізми активації такої деградації не з’ясовані, але швидке руйнування транскрипту має стимулювати дисоціацію полімеразного комплексу. Термінація транскрипції іноді потребує також інших регуляторних елементів послідовності. Одні з них можуть знаходитись у межах останнього інтрона та стимулювати / блокувати впізнання polyA-сигналу, інші – нижче від polyA-сигналу й зумовлювати паузи в роботі полімерази, які сприяють термінації.
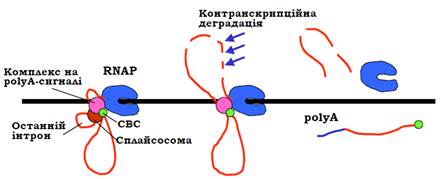
Рис. 9. Синхронізація поліаденілування і термінації транскрипції.
Узагальнену схему будови зрілої еукаріотичної мРНК, яка звільняється з полімеразного комплексу, транспортується в цитоплазму і використовується як матриця для білкового синтезу, зображено на рис. 10. Між кепом і початком кодуючої ділянки (стартовим кодоном – найчастіше AUG) розташована 5′-кінцева зона, що не транслюється (5′ UTR – UnTranslated Region). За кодуючою ділянкою, що закінчується одним із стопкодонів, і перед polyA послідовністю розташована 3′-кінцева зона, що не піддається трансляції. Обидві зони, що не транслюються, містять важливі елементи послідовності, які використовуються для регуляції білкового синтезу.
Майже єдиним винятком серед інших еукаріотичних мРНК є гістонові мРНК, які не піддаються сплайсингу (гістонові гени не містять інтронів) і поліаденілуванню 3′-кінця. Термінація транскрипції гістонових генів залежить від двох сигнальних елементів послідовності РНК: один утворює шпильку (як у прокаріотів), інший спарюється з маленькою ядерною РНК U7; між цими елементами індукується розріз. Аналогічно відбувається термінація транскрипції маленьких ядерних РНК, які також не піддаються поліаденілуванню.

Рис. 10. Схема будови мРНК.
Дата добавления: 2015-09-11; просмотров: 1424;