Альтернативний сплайсинг
ПремРНК, що синтезується під час транскрипції, може піддаватися сплайсингу та поліаденілуванню різними альтернативними шляхами: кілька екзонів на початку чи всередині гена можуть вирізатися з транскрипту, останній може обрізатися та піддаватися поліаденілуванню за рахунок використання polyA-сигналу всередині одного з інтронів тощо. У результаті утворюються різні молекули мРНК, що містять різні набори екзонів і, відповідно, кодують різні-білки. Багатокомпонентність (і необхідність кооперації між компонентами) сплайсосоми та системи розрізання / поліаденілування дозволяє здійснювати тонку регуляцію утворення мРНК певного типу за рахунок зміни транскрипційної активності генів, що кодують тічи інші компоненти машинерії процесингу, зміни концентрацій компонентів, їхньої хімічної модифікації.
Ключова роль у визначенні шляху сплайсингу належить білкам-регуляторам сплайсингу (SR). Досить часто регулятор специфічно зв’язується з певною послідовністю нуклеотидів усередині екзоната стимулює впізнання сплайс сайтів по обидва боки від нього(рис. 11, а). Зрозуміло, що в разі відсутності регулятора такий екзон буде вирізано разом з інтронами, що його фланкують.

Рис. 11. Кілька варіантів участі сплайсинг-регуляторів, що зв’язуються всередині екзона (а, б, в) або інтрона (г, д, е), у визначенні шляху сплайсингу.
У випадку негативної регуляції зв’язування регулятора утруднює впізнання сплайс сайтів. Наприклад, взаємодія з регулятором ініціюється всередині екзона, молекули білка кооперативно зв’язуються поруч одназ одною і врешті решт блокують 3′-сплайс сайт (рис. 11, б). Замість заблокованого сприймається наступний такий сайт, і екзон вирізається з молекули разом із двома інтронами, що його фланкують. За наявності іншого регулятора, який також упізнає специфічну послідовність в екзоні, процес зростання білкового комплексу зупиняється, і 3′-сплайс сайт стає вільним для впізнання елементами сплайсосоми (рис. 11, в).
Сплайсинг-регулятори можуть також мати сайти зв’язування в інтронах, впливаючи на ефективність збирання сплайсосоми (рис. 11, г). Регулятор може блокувати, наприклад, 3′-сплайс сайт: якщо така блокада відсутня, сайт розпізнається (рис. 11, д), за наявності регулятора сприймається інший 3′-сплайс сайт усередині екзона – у складі мРНК залишається скорочений екзон (рис. 11, е). Вибір шляху утворення кінцевого мРНК-продукту часто залежить також від альтернативного вибору polyA-сигналів, які можуть бути присутніми не лише нижче останнього екзона, а й усередині інтронів.
Приклад такого типу зображено на рис. 12. Узагалі, якщо polyA-сигнал міститься всередині інтрона, його впізнання блокується 5′-сплайс сайтом – асоційованими з U1 білками (polyA-сигнал, що розташований після термінального екзона, не блокується, оскільки немає 5′-сплайс сайта). У результаті такої блокади polyA-сигнал не впізнається і вирізається разом з інтроном. Але якщо діє специфічний сплайсинг-регулятор, який заважає впізнанню 5′-сплайс сайта або взаємодії U1 білків з елементами системи впізнання polyA-сигналу, сигналу пізнається, і мРНК втрачає всі екзони, розташовані нижче від сигналу. Наприклад, сплайсинг мРНК важкого ланцюга імуноглобуліну IgM спрямовується першим шляхом (внутрішній polyA-сигнал ігнорується) у В-лімфоцитах, що розвиваються, – синтезується мембранна форма білка, яка містить С-кінцевий домен, зв’язаний з мембраною. У плазматичних клітинах спрацьовує внутрішній polyA-сигнал (упізнання сигналу підсилюється також завдяки зростання концентрації CstF), і синтезується секреторна форма білка.
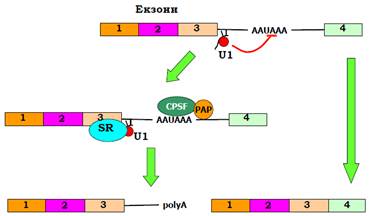
Рис. 12. Приклад двох шляхів процесингу, вибір яких залежить від впізнання polyA-сигналу всередині інтрона.
Поряд із концентрацією елементів машинерії процесингу та сплайсинг-регуляторами, досить важливим фактором регуляції сплайсингу є швидкість транскрипції (як у розумінні швидкості руху полімерази, так і в сенсі кількості полімераз, що одночасно працюють на даному гені).
У випадку інтрона середньої довжини 3′-сплайс сайт синтезується РНК-полімеразою через 15 – 30 хвилин після 5′-сплайс сайта. За цей час упізнання 5′-сплайс сайта вже завершилося, і перший 3′-сплайс сайт упізнається як такий. Якщо за досить короткий час синтезується кілька 3′-сплайс сайтів, виникає конкуренція між ними за відповідні елементи сплайсосоми. При цьому перший із сайтів буде вигравати при низькій швидкості руху полімерази (рис. 13).
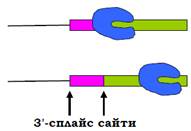
Рис. 13. Різні положення РНК-полімерази через однаковий проміжок часу внаслідок різниці у швидкості транскрипції.
Швидкість транскрипції в розумінні кількості полімераз, що одночасно працюють на даному гені, також впливає на процесинг: чим нижчою є швидкість, тим більше часу розібратися із сигналами, на великій швидкості полімераза може проскочити через екзон чи polyA-сигнал. При цьому слід зауважити, що швидкість (частота рекрутування полімераз до промотора) визначається на стадії ініціації та контролюється транскрипційними факторами. Отже, значною мірою шлях процесингу визначається вже на етапі ініціації транскрипції. При цьому до промотора (відповідно, і до РНК-полімеразного комплексу) можуть рекрутуватися також і регулятори сплайсингу. Тому на химерних генах із чужорідним промотором часто виявляються дефекти процесингу.
Дата добавления: 2015-09-11; просмотров: 1851;