Сплайсосома: механізм сплайсингу
Обидві реакції сплайсингу здійснюються у складі спеціальної мультимолекулярної структури – сплайсосоми (spliceosome), яка зв’язана з CTD і утворюється на кожному інтроні за участю самої премРНК, білків і особливих молекул маленьких ядерних РНК (small nuclear RNA, snRNA).
Маленькі ядерні РНК, які синтезуються РНК-полімеразою ІІ на відповідних генах, згрупованих у кластери, відіграють ключову роль у визначенні просторової структури, формуванні та функціонуванні сплайсосоми. У сплайсингу беруть участь п’ять типів маленьких ядерних РНК (довжиною 100 – 200 нуклеотидів): U1, U2, U4, U5 та U6. Маленькі ядерні РНК, які існують у клітині у вигляді комплексів зі специфічними білками, мають певну просторову структуру за рахунок утворення комплементарних подвійних спіралей – шпильок. Завдяки комплементарному спарюванню з консенсусними послідовностями інтрона відбувається їхня взаємодія з премРНК та іншими маленькими ядерними РНК. Наприклад, РНК U2 впізнає консенсус у зоні розгалуження, причому згаданий аденіновий нуклеотид залишається неспареним – випетльовуюється між сусідніми парами основ (рис. 5).
Саме ця обставина визначає його особливий конформаційно-напружений стан і, як наслідок, підвищену реакційну здатність. Оскільки подвійні спіралі РНК у сплайсосомі короткі, вони потребують додаткової стабілізації. Крім того, під час збирання та функціонування сплайсосоми певні спіралі мають бути зруйновані й замінені на інші. Обидві операції забезпечуються білками сплайсосоми. Отже, роль білків сплайсосоми зводиться до:
• розкручування подвійних спіралей РНК АТР-залежними геліказами;
• стабілізації подвійних спіралей і загальної просторової структури сплайсосоми білками зі специфічною спорідненістю до РНК;
• регуляції сплайсингу – блокування чи підсилення ефективності збирання сплайсосоми на даному інтроні білками-регуляторами сплайсингу (SR, Splicing Regulators).
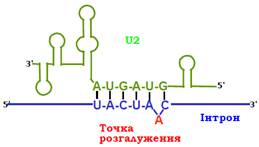
Рис. 5. Схема структури маленької ядерної РНК U2 у комплексі з зоною розгалуження інтрона.
Загальний сценарій збирання та роботи сплайсосоми схематично зображено на рис. 6. Після синтезу 5′-кінцевої зони інтрона, вона впізнається маленькою ядерною РНК U1 за рахунок комплементарного спарювання між U1 і 5′-сплайс сайтом (U1 при цьому зв’язана з відповідними білками). Далі в міру синтезу премРНК зона розгалуження впізнається специфічним білком ВВР (Branch point Binding Protein, позначається також як SF1 у ссавців). 3′-кінцева зона інтрона та Y-збагачений трек упізнаються допоміжним білком U2АF (U2 Auxiliary Factor), який далі сприяє заміні ВВР на РНК U2. Зв’язування U2 (яке, власне, визначає точку розгалуження, див. рис. 5) потребує АТР-залежного руйнування певних подвійних спіралей у складі цієї молекули. На наступних стадіях збирання та перебудов сплайсосоми (рис. 6) також відбувається АТР-залежне руйнування частини подвійних спіралей і заміна їх іншими.
Після зв’язування U2, на завершальному етапі збирання, з інтроном взаємодіє потрійний комплекс U4U5U6: U4 при цьому звільнюється, а U6 витісняє U1, взаємодіючи з 5′-кінцевою зоною інтрона. Крім того, U6 взаємодіє з U2, сприяючи наближенню 5′-сплайс сайта до точки розгалуження. Додатково структура стабілізується маленькою ядерною РНК U5, яка взаємодіє з 3′-кінцем першого екзона та іншими елементами сплайсосоми. Після формування сплайсосоми здійснюється її структурна перебудова (АТР-залежне розплітання частини подвійних спіралей геліказами), результатом якої є безпосереднє наближення 5′-сплайс сайта до точки розгалуження: виникають умови для першої реакції транс-естерифікації з утворенням ласо (див. також рис. 4). Хімічна перебудова інтрона викликає нову перебудову просторової структури сплайсосоми: за рахунок одночасної взаємодії з U5 наближуються один до одного кінці екзонів, що створює умови для другої транс-естерифікації.

Рис. 6. Схема збирання сплайсосоми та її функціонування.
На останньому етапі від двох уже з’єднаних екзонів АТР-залежним шляхом (порушення комплементарних взаємодій) видаляється комплекс інтрона з маленькими ядерними РНК. Отже, сплайсосома працює як АТР-залежна молекулярна машина.
Структурні перебудови машини забезпечують перегрупування елементів – субстратів реакцій сплайсингу. Каталіз обох реакцій сплайсингу здійснюється молекулами РНК. Механізм каталізу є таким самим, як для білкових ферментів. Наприклад, для першої реакції активний центр формується зоною розгалуження інтрона й маленькими ядерними РНК U2 та U6. Просторова структура активного центру жорстко утримує субстрати – 5′-сплайс сайт і точку розгалуження – у певній взаємній орієнтації та забезпечує підвищену реакційну здатність аденінового нуклеотиду, який можна розглядати одночасно і як субстрат у збудженому проміжному стані, і як компонент активного центру. У хлоропластах сплайсинг відбувається за механізмом так званого самосплайсингу (selfsplicing), тобто без участі білків і маленьких РНК: мРНК сама набуває просторової структури, яка має каталітичну активність щодо власного сплайсингу.
За аналогією з білковими ензимами, молекули РНК, які мають каталітичну активність, називають рибозимами. Одним із таких рибозимів є сплайсосома. Зазвичай каталітична активність рибозима (як і у випадку сплайсосоми) залежить від білків, які виконують допоміжні функції, стабілізуючи структуру активного центру. Вважається, що на ранніх етапах добіологічної еволюції головними (або єдиними) біологічними макромолекулами були молекули РНК (так званий РНК-світ),оскільки це єдиний тип макромолекул, що можуть одночасно виконувати роль носіїв спадкової інформації та виступати каталізаторами.
Пізніше більш стабільні молекули ДНК перебрали на себе роль носіїв інформації, а різноманітніші за просторовою структурою білки – роль каталізаторів. Але в кількох важливих випадках (і сплайсинг – не єдиний приклад) рибозими виявилися еволюційно консервативними. Сплайсосоми утворюються на інтронах премРНК (і на CTD РНК-полімерази) послідовно під час транскрипції – майже відразу після синтезу сплайс-сайтів вони впізнаються відповідними елементами (рис. 7). При цьому СВС, що зв’язаний з кепом, підсилює ефективність зв’язування U1 у першому 5′-сплайс сайті – напевно, шляхом прямої взаємодії; вплив СВС на сплайсинг наступних інтронів послаблюється.
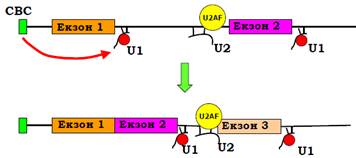
Рис. 7. Послідовне збирання сплайсосом на інтронах під час транскрипції.
Швидкість збирання сплайсосом визначається швидкістю транскрипції. З іншого боку, зі сплайсосомою взаємодіють фактори елонгації транскрипції – наявність сплайс-сайта сприяє прискоренню руху полімерази. Такий процес синтезу РНК та її сплайсингу продовжується до того моменту, поки у складі преРНК не з’являється специфічна послідовність – сигнал термінації.
Дата добавления: 2015-09-11; просмотров: 1525;