Роль білків HMG в активації транскрипції
Назва HMG об’єднує три, структурно абсолютно різні, родини білків. Але всі вони задіяні до системи регуляції транскрипції в еукаріотів.
HMGA – невпорядковані поліпептиди, що містять у своєму складі три так звані АТ-гаки – елементи амінокислотної послідовності, здатні впізнавати короткі (чотири – п’ять пар основ), часто присутні в промоторах, АТ-збагачені ділянки ДНК. Невпорядковані частини поліпептидного ланцюга, які з’єднують АТ-гаки, мають спорідненість до певних транскрипційних факторів. Отже, зв’язування HMGA може ініціювати збирання енхансосоми. Гени HMGA активуються в результаті численних шляхів сигнальної трансдукції.
Один із прикладів участі HMGA в активації транскрипції наведено на рис. 19.
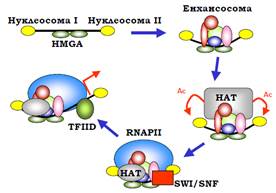
Рис. 6.19. Послідовність подій при активації β-інтерферонового промотора.
HMGA зв’язується в лінкері між двома позиційованими нуклеосомами у промоторі β-інтерферонового гена. На цьому зародковому елементі відбувається збирання енхансосоми, до складу якої рекрутується гістонацетилтрансферазний комплекс. Ацетильовані Lys упізнаються іншою НАТ, яка продовжує процес ацетилювання, а також відбувається збирання преініціаторного комплексу – включаючи РНК-полімеразу ІІ, але за винятком TFIID. Ацетильовані Lys та інші компоненти енхансосоми рекрутують SWI/SNF, який змінює позицію нуклеосоми ІІ (рис. 19), що закривала ТАТА-бокс. Унаслідок цього нарешті зв’язується TFIID, і запускається транскрипція.
Білки HMGA не обов’язково виступають активаторами транскрипції – вони можуть також блокувати сайти зв’язування транскрипційних факторів, сприяючи репресії певних промоторів.
HMGВ – невеликі глобулярні білки, що містять один (дріжджі, комахи, рослини) або два (хребетні) HMG-бокси. У клітинних ядрах ссавців HMGВ присутні у значній кількості – до 10 – 20 копій на одну нуклеосому. Не дуже міцне (на невеликий проміжок часу) зв’язування HMG-боксу з маленьким жолобком ДНК індукує значний вигин на ~80°. Відповідно, білки HMGВ є загальними лабілізаторами структури хроматину (їх називають також архітектурними білками або хроматиновими шаперонами), що діють за принципом торкнувся й пішов. Після зв’язування та дисоціації HMGВ може нічого не відбутися. Але вигин, який індукується HMG-боксом може або підвищити спорідненість транскрипційних факторів, або додатково стабілізувати структуру енхансосоми (рис. 20). Далі HMGВ може або залишитись у складі енхансосоми, або дисоціювати, замінившись на інші транскрипційні фактори.
Крім того, білки HMGВ є конкурентами гістону Н1: зв’язування їх із ДНК на вході до нуклеосоми полегшує тимчасову деконденсацію хроматинової фібрили. Завдяки зв’язуванню з ДНК на вході до нуклеосоми HMGВ виступає також як кофактор комплексів ремоделювання родини Swi/Snf: вигин, що індукується HMG-боксом, полегшує випетлювання ДНК на поверхні нуклеосоми, яке мігрує всередину нуклеосомної ДНК (рис. 21). Ремоделюванню за таким механізмом сприяє не обов’язково білок HMGВ – HMG-бокси часто містяться також у складі самих комплексів ремоделювання.
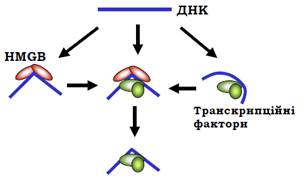
Рис. 20. Залучення білків HMGВ до збирання енхансосоми.
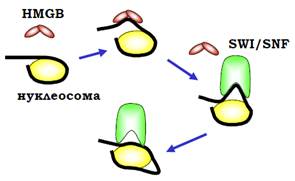
Рис. 21. Залучення білків HMGВ до ремоделювання хроматину.
HMGN – невпорядковані поліпептиди довжиною близько 100 амінокислот. За рахунок позитивно зарядженої частини ланцюга взаємодіють з ДНК всередині нуклеосоми. Негативно заряджена частина молекули конкурує з ДНК за гістонові хвости. У результаті HMGN, який накопичується у транскрипційно активних ділянках, сприяє деконденсації хроматинової фібрили. Взаємодія HMGN з нуклеосомою – слабка, тобто він, як і HMGВ, лише на короткий час підвищує доступність ДНК для інших елементів системи активації транскрипції.
Дата добавления: 2015-09-11; просмотров: 1087;