Розміри геномів і кількість білкових генів деяких організмів.
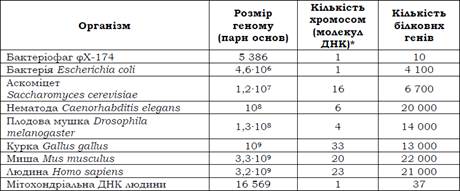
Прокаріотичні й вірусні геноми. Схему організації одного з найпростіших (і першого з вивчених) геномів – геному бактеріофага φХ-174 (слід читати фі-десять) – показано на рис. 3. Геном побудований надзвичайно економно: десять генів (один із них дає два різні транскрипти) займають практично всю циркулярну ДНК бактеріофага. Більше того, декілька генів перекриваються за рахунок використання різних рамок зчитування: гени А і С та С і D перекриваються своїми кінцями, гени B, K і E повністю знаходяться у межах інших генів; три гени – А, С і K – використовують усі три можливі рамки зчитування на одній ділянці ДНК (звичайно, у даному випадку всі три рамки є відкритими). Явище перекриття генів за рахунок використання різних рамок зчитування спостерігається також для кількох інших бактеріофагів, проте іноді зустрічається в еукаріотів.
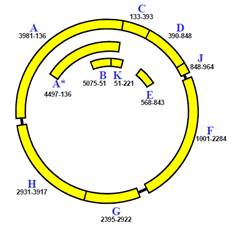
Рис. 3. Геном бактеріофага φХ-174. Позначено початок і кінець кожного гена, загальна
довжина ДНК – 5386 пар основ.
Загалом геноми вірусів і бактеріофагів побудовані за подібним економним принципом: порівняно невелика кількість генів, мінімальна кількість міжгенної ДНК.
У геномі прокаріотичної клітини кількість ДНК і генів значно зростає, але зберігається принцип економічності щодо використання більшості послідовностей для кодування генетичної інформації. Наприклад, геном Escherichia coli представлений однією циркулярною молекулою ДНК (бактеріальною хромосомою) довжиною 4,6 млн пар основ. Близько 90 % цієї ДНК припадає на кодуючі послідовності ~4100 білкових генів і ~120 генів РНК, що не транслюється. Середній розмір гена складається з 950 пар основ, середня довжина міжгенної ділянки – 118 пар основ. Проте, міжгенні зони мають досить нерівномірний розподіл за довжиною, яка варіює від 0 до 1 тис. 730 пар основ.
Приблизно ¾ транскрипційних одиниць E. coli містять один ген, решта реалізує характерний для бактерій (і тільки для них) оперонний принцип організації генетичного матеріалу. Оперон являє собою кластер так званих структурних генів, на яких синтезується одна молекула мРНК, що має кілька (на кожен структурний ген) послідовних (які не перекриваються) відкритих рамок зчитування для трансляції відповідних білків. У межах оперона згруповані структурні гени, які відповідають за синтез білків, залучених до одного ланцюжка біохімічних перетворень (ферменти синтезу або деградації певної сполуки). Крім структурних генів оперон має регуляторні ділянки, за рахунок яких здійснюється регуляція транскрипції оперона як цілого. У геномі E.coli міститься ~650 таких одиниць транскрипції.
Характерною особливістю бактерій є наявність у клітині, поряд із бактеріальною хромосомою, невеликих автономних елементів геному – плазмід. Плазміда є циркулярною молекулою ДНК (типовий розмір ~3 тис. пар основ), яка містить кілька генів і реплікується незалежно від бактеріальної хромосоми. Плазміди широко використовуються як зручний інструмент молекулярно-біологічних досліджень.
Циркулярна ДНК бактеріальної хромосоми існує у клітині у вигляді комплексу з білками. Ця взаємодія є досить динамічною, і практично весь бактеріальний геном (що принципово відрізняє його від еукаріотичного геному, див. нижче) перебуває в потенційно транскрипційно-активному стані: гени й оперони є об’єктами швидкої оперативної регуляції у відповідь на зміну зовнішніх умов.
Еукаріотичні геноми містять значно більшу кількість ДНК порівняно з геномами прокаріотів (див. табл. 1), причому переважна частина цієї ДНК представлена послідовностями, що не є кодуючими. У тому числі приблизно половина еукаріотичного геному – це послідовності, представлені багатьма копіями (послідовності, що повторюються). Еукаріотична ДНК існує у клітинному ядрі у складі хромосом, кожна хромосома містить одну гігантську лінійну молекулу ДНК. Послідовності, що повторюються, зосереджені на кінцях хромосом (теломери) і в зонах прикріплення хромосом до веретена поділу при мітозі (центромери).
Характерною ознакою генів еукаріотів (на відміну від прокаріотів) є мозаїчний принцип будови кодуючої частини: власне кодуюча частина – це послідовність окремих змістовних ділянок – екзонів (exon), розділених беззмістовними інтронами (intron). Часто екзони відповідають окремим структурним доменам мультидоменних білків: еволюційне збирання білка з кубиків доменів може здійснюватись шляхом перетасування екзонів на рівні ДНК. Беззмістовними інтрони є в тому сенсі, що не містять інформації про кінцевий продукт, але в межах інтронів часто розташовані важливі регуляторні ділянки. При транскрипції молекула РНК синтезується суцільно (первинний продукт транскрипції – первинний транскрипт – містить екзони та інтрони).
Отже, необхідним етапом експресії гена є процес сплайсингу – вирізання інтронів і зшивання екзонів у кінцевий транскрипт, який може бути використаний як матриця для білкового синтезу. При цьому сплайсинг може бути спрямований різними шляхами (рис. 4) – альтернативний сплайсинг, – унаслідок чого утворюються різні кінцеві продукти – різні білки. Таким чином, альтернативний сплайсинг можна інтерпретувати як механізм перекриття еукаріотичних генів.
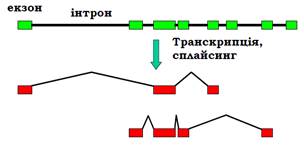
Рис. 4. Мозаїчна будова кодуючої частини гена та схема утворення різних кінцевих продуктів унаслідок альтернативного сплайсингу.
Інший випадок перекриття генів у геномах еукаріотів пов’язаний з можливістю розташування гена (зі своїми інтронами та екзонами) у межах інтрона іншого гена. В окремих випадках спостерігається також перекриття відкритих рамок зчитування.
Усі гени багатоклітинного організму можна розділити на дві групи:1) гени, від яких залежать певні універсальні функції та які активні в усіх клітинах, – “гени домашнього господарства” (hous ekeeping genes); 2) гени, що специфічно активуються у клітинах певного типу, – гени розкоші– (luxury genes). Загальною ознакою генів першої групи є розташування в їхніх регуляторних зонах CpG-острівців (CpG-islands) – ділянок із підвищеним вмістом динуклеотидів CpG (контактів типу CG між парами основ). Вміст цих динуклетидів в еукаріотичних геномах приблизно у п’ять разів менший за очікуваний унаслідок метилування цитозину в складі CpG-контакту: 5mC (5-метилцитозин) спонтанно перетворюється на тимін. Метилування цитозину в регуляторних ділянках є одним із механізмів репресії генів. Відповідно, гени, що зберігають свою активність у більшості клітин, містять неметильовані динуклеотиди CpG, вміст яких зберігається на високому рівні (для ДНК випадкової послідовності із 40 % GC-пар очікуваний вміст динуклеотидів CpG становить 4 %). Типове значення вмісту GC-пар в еукаріотичних геномах – близько 40 % (у геномі людини – 41 %). Проте спостерігаються варіації локального GC-вмісту в широких межах – від 30 до 60 %; зустрічаються й досить довгі ділянки (до кількох мільйонів пар основ) з аномальним вмістом GC-пар. Загальна тенденція при цьому така: високий вміст GC-пар корелює з високою щільністю генів, зменшеною довжиною інтронів і низькою щільністю послідовностей, що повторюються.
Серед еукаріотичних генів 25 – 50 % є унікальними (представлені в геномі єдиною копією), решта належать до родин генів, що складаються з кількох копій, як правило, не ідентичних. Відповідні (гомологічні але не ідентичні) білки складають родину білків. Кілька родин (протеїнкінази, транскрипційні фактори певного типу, імуноглобуліни) містять сотні білків, більшість родин складається з кількох (до 30) білків. Гени такої родини часто об’єднані в геномі в кластери – розташовані поряд у певній хромосомі (кластери генів теплового шоку, глобінові гени). Слід зауважити, що такий кластер не є опероном – кожен ген піддається регуляції як окрема одиниця транскрипції. Наприклад, кластер генів β-субодиниці гемоглобіну містить гомологічні гени, які активуються на певних стадіях індивідуального розвитку (рис. 5).
Рис. 5. Кластер β-глобіну в 16-й хромосомі людини (кожен ген містить інтрони).
Указано стадії розвитку, на яких відповідні гени є активними.
β-Глобіновий кластер містить також неактивний псевдоген. Псевдогени – це послідовності, гомологічні певним генам, але не експресуються. До такої дезактивації гена може призвести мутація, яка порушує ініціацію транскрипції, сплайсинг, викликає невчасну термінацію тощо. Після дезактивації псевдоген перестає бути об’єктом відбору, і в ньому накопичується велика кількість мутацій. Зрозуміло, що в першу чергу псевдогени виникають саме в кластерах – коли є кілька копій гена, і пошкодження одного з них не приводить до фатальних наслідків. Типовою ознакою еукаріотичних геномів є наявність кількох типів генних кластерів, що повторюються багато разів. Серед білкових генів це стосується генів гістонів – структурних білків хроматину. Гени п’яти молекул гістонів завжди згруповані в кластер (кожен ген – окрема одиниця транскрипції), який повторюється до 100 разів. Іншим прикладом кластерів, що повторюються, є гени рибосомної РНК (рис. 6, тип рибосомних РНК позначають за їхньою константою седиментації).
У цьому випадку кластер є одиницею транскрипції, первинний транскрипт піддається далі дозріванню – частковій деградації з утворенням окремих молекул рРНК. Такий тандемний повтор рибосомного кластера є ядерцевим організатором – зоною, де утворюється ядерце (місце збирання рибосом). Гени рРНК ще одного типу – 5S – також тандемно повторюються в іншому місці геному. Також зібрані в тандемні кластери частина генів тРНК і маленьких ядерних РНК.
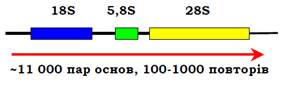
Рис. 6. Кластер генів рибосомних РНК, який повторюється в зоні ядерцевого організатора. Червона стрілка позначає первинний транскрипт.
Загальна кількість генів у геномах вищих еукаріотів варіює приблизно від 20 до 30 тис. (табл. 1). Так, у геномі людини кількість білкових генів становить 21 тис., ще кілька тисяч генів кодують РНК, яка не піддається трансляції. Наведемо як приклад кілька загальних характеристик білкових генів людини:
• Середній розмір гена (ураховуючи інтрони) – 100 тис. пар основ (максимум – 2 млн 400·тис.).
• Середній розмір кодуючої частини – 1 тис. 400 пар основ (максимум – 80 тис.).
• Середній розмір екзона – 150 пар основ.
• Середня кількість екзонів – 9 (максимум – 178).
• Середній розмір інтрона – 5 тис. пар основ (максимум – 820 тис.).
Загальною тенденцією, що спостерігається в геномі людини, є наявність порівняно коротких екзонів, розділених довгими інтронами. Приблизний розподіл білків людини (сукупність усіх білків організму називається протеомом) за їхніми функціями зображено на рис. 7. Як видно з рис. 8, кодуючі послідовності цих генів займають лише ~1,5 % геному. Решта припадають на міжгенну ДНК (де містяться також регуляторні ділянки), інтрони (~34 %) і, більше половини геному, – на послідовності, що повторюються.
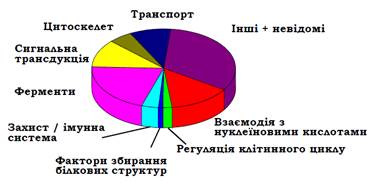
Рис. 7. Розподіл білків протеому людини за функціями.

Рис. 8. Відносний вміст послідовностей різних типів у геномі людини.
Основні типи повторів, присутні в геномі вищих еукаріотів:
1. Псевдогени – від 1 до кількох тисяч у геномі людини.
2. Тандемні повтори. До цього класу повторів можна віднести тандемні повтори генних кластерів, які вже згадувалися вище. Крім того, до тандемних повторів відносять багатократні повтори коротких послідовностей по шість–вісім пар основ у теломерах (TTAGGG у людини) і повтори α-сателітної ДНК у центромерах (довжина повтору варіює від 7 пар основ у дрозофіли до 200 пар основ у ссавців, у людини – 171 пара основ). По всьому геному розподілені також так звані прості повтори (SSR, simple sequence repeats, ~3 % у геномі людини). Зазвичай виділяють мікросателіти – 1 – 15 пар основ, що повторюються від 10 до кількох тисяч разів, і мінісателіти – 15 – 500 пар основ, які повторюються до 100 разів. У геномі людини є принаймні 30 тис. міні- та 200 тис. мікросателітних локусів.
3. Сегментні дуплікації – великі блоки довжиною 1 – 200 тис. пар основ, які характеризуються високим ступенем гомології (близько5 % у геномі людини). Імовірно, сегментні дуплікації є продуктом порушення хромосом. Частіше зустрічаються в перицентромерних і субтеломерних зонах.
4. Інтерсперсні (мобільні) елементи, здатні до переміщення та розмноження в межах геному (~44 % у геномі людини). Значна частина таких послідовностей є результатом колишньої активності мобільних елементів (таких, що втратили здатність до переміщення), але деякі зберігають свою активність досі. Основні типи мобільних елементів у геномі людини:
• ДНК-транспозони (3 %) – переміщення здійснюється шляхом вирізання ділянки ДНК із наступним вбудовуванням її в інше місце.
• LTR-ретропозони (8 %) – як і для інших двох типів мобільних елементів, переміщення відбувається через проміжну молекулу РНК: на елементі послідовності (який містить довгі кінцеві повтори – Long Terminal Repeats – і кілька генів, зокрема ген зворотної транскриптази) здійснюється транскрипція; молекула РНК прямує до цитоплазми, де відбувається зворотна транскрипція – синтез ДНК на РНК матриці за допомогою зворотної транскриптази (РНК-залежна ДНК-полімераза). ДНК-повертається до ядра, де вбудовується в геном.
• Мобільні елементи LINE (Long INterspersed Elements, 20 %) містять кілька генів, включаючи ген зворотної транскриптази. На відміну від LTR-ретропозонів, зворотна транскрипція здійснюється в ядрі. У геномі людини присутні три родини таких елементів, одна з них є активною і слугує основним джерелом зворотної транскриптази у клітині.
• Мобільні елементи SINE (Short INterspersed Elements, 13 %) – короткі (100 – 400 пар основ) беззмістовні елементи, які використовують для переміщення систему LINE. До цього класу належить і Alu-повтор (від назви відповідної рестриктази, яка здатна специфічно гідролізувати цей елемент послідовності).
Мобільні елементи розподілені в геномі нерівномірно: є довгі ділянки, що на 90 % складаються з цих елементів, і такі, де інтерсперсні елементи відсутні. Загалом спостерігається негативна кореляція між щільністю генів і мобільних елементів. Винятком із цієї закономірностіє позитивна кореляція між щільністю генів і елементів типу SINE.
Геном мітохондрій і хлоропластів – автономний елемент еукаріотичного геному. Циркулярна (як правило) молекула мітохондріальної ДНК містить від 6 тис. до 2 млн. пар основ і певний набір генів (рРНК, тРНК, деяких білків). Мітохондріальні гени містять інтрони (але не у ссавців). Розмір мітохондріального геному знижується в ході еволюції. Циркулярна ДНК хлоропластів більша за розміром – містить до 200 тис. пар основ, хлоропластний геном кодує до ~100 білків.
Дата добавления: 2015-09-11; просмотров: 8931;