Нуклеосома
Білковий компонент нуклеосоми – гістони, які є одним з найбільш еволюційно консервативних класів білків. Усі корові гістони (містять від 102 до 135 амінокислотних залишків) мають спільну схему будови. У первинній структурі виділяють дві частини: глобулярну та N кінцеву невпорядковану (хвіст) довжиною від 20 (Н2А) до 40 (Н3) амінокислотних залишків. Гістон Н2А має також помітний С-кінцевий хвіст довжиною близько 15 залишків. Невпорядковані хвости практично не містять гідрофобних залишків, збагачені позитивно зарядженими амінокислотами і є субстратами для численних посттрансляційних модифікацій (про що йтиметься нижче).
Глобулярна частина всіх корових гістонів, у свою чергу, також має спільну структуру. Вона виглядає як характерний триспіральний гістоновий мотив (histone fold), у якому одна довга α-спіраль фланкована двома короткими. Гістон Н3 містить також додаткову α-спіраль з боку N-кінця мотиву (α-N-спіраль), а гістон Н2В – додаткову α-С-спіраль. Гістоновий мотив не формує гідрофобного ядра, заекранованого від розчинника, – значна кількість гідрофобних залишків опиняється на поверхні. Унаслідок цього одна молекула корового гістону не може існувати як окремий глобулярний білок у водному середовищі. Мінімальними стабільними структурними одиницями є гетеродимери Н2АН2В та Н3Н4 (мають подібну структуру). Два гістонові мотиви формують у складі димерів щільне гідрофобне ядро, а специфічність формування димерів залежить від наявності додаткових α-N- та α-С-спіралей у гістонах Н3 та Н2В відповідно.
На поверхні димеру розташовані три зони скупчення позитивно заряджених амінокислотних залишків, які, відповідно до електростатичного механізму, здатні взаємодіяти з ДНК. У межах кожного такого сайта є принаймні один залишок аргініну, який відіграє найважливішу роль у взаємодії з ДНК. Разом ці три сайти створюють платформу для зв’язування ділянки ДНК довжиною 27 – 28 пар основ (~ 2,5 витка подвійної спіралі,).
Два гетеродимери Н3Н4 взаємодіють між собою за рахунок утворення чотириспірального пучка між гістоновими мотивами двох молекул Н3. У результаті формується тетрамер (Н3Н4)2 – центральний комплекс у структурі нуклеосоми (рис. 9). Структура тетрамеру нагадує підкову, яка характеризується хіральністю – утворює елемент лівої спіралі. Вісь симетрії тетрамерного комплексу (яка водночасє і віссю симетрії всієї нуклеосоми) проходить через інтерфейс між двома молекулами Н3.
Аналогічний за своєю структурою чотириспіральний пучок між гістоновими мотивами молекул Н4 та Н2В забезпечує взаємодію між тетрамером (Н3Н4)2 і димером Н2АН2В. Таким чином, підкова тетрамеру симетрично продовжується двома димерами Н2АН2В в обидва боки, результатом чого є утворення октамеру (Н2АН2ВН3Н4)2. Отже, глобулярні частини гістонів утворюють октамерний комплекс, який і служить білковим кором нуклеосоми (рис. 10). На глобулярній поверхні октамеру існує своєрідний трек позитивно заряджених амінокислотних залишків, який використовується для взаємодії з нуклеосомною ДНК довжиною 145 пар основ.
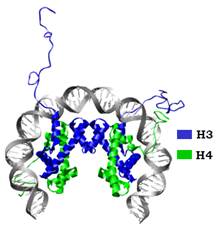
Рис. 9. Тетрамер гістонів (Н3Н4)2 у комплексі з ДНК у складі нуклеосоми (1KX5).
Окрема нуклеосома (нуклеосомна кор-частинка) – октамер гістонів + 145 пар основ ДНК – може бути вилучена з хроматину за допомогою мікрококової нуклеази: вона робить дволанцюговий розріз у ДНК, а оскільки один із ланцюгів нуклеосомної ДНК завжди взаємодіє з гістонами (рис. 10), доступною для нуклеази є лише ДНК за межами нуклеосоми. У хроматині вся ДНК формує нуклеосоми із середньою щільністю одна нуклеосома на 200 пар основ, сусідні нуклеосоми з’єднані міжнуклеосомними лінкерними (linker) ділянками. Нуклеосомна ДНК разом з лінкерною ділянкою складають так званий нуклеосомний повтор, довжина якого (середнє значення 200 пар основ) варіює як уздовж полінуклеосомного ланцюга, так і залежно від функціонального стану, типу клітин тощо.
Вісь симетрії нуклеосоми проходить через центральну точку нуклеосомної ДНК, у якій великий жолобок подвійної спіралі є зверненим до поверхні октамеру гістонів (рис. 10). ДНК-гістонові взаємодії здійснюються в позиціях, де маленький жолобок контактує з поверхнею октамеру: саме тут реалізуються взаємодії ДНК із позитивно зарядженими сайтами на поверхні гістонових мотивів. Як видно з рис. 10, нуклеосомна ДНК є значно вигнутою на поверхні октамеру гістонів і формує майже ідеальну ліву суперспіраль із радіусом 41,9 Å і кроком 25,9 Å. Центральні 133 пари основ такої суперспіралі дають 1,67 витка, решта нуклеосомної ДНК (дві ділянки по сім пар основ на вході/виході) є практично прямою і просто продовжує хід нуклеосомної суперспіралі. Зрозуміло, що значний вигин нуклеосомної ДНК потребує енергетичних витрат, які компенсуються ДНК-гістоновими взаємодіями.
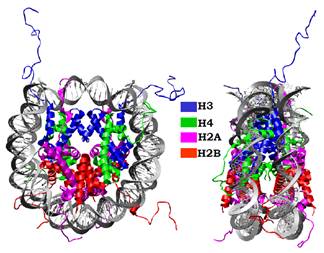
Рис. 10. Структура нуклеосоми у двох проекціях (1KX5).
Структура нуклеосомної ДНК зумовлена її взаємодією із глобулярною частиною октамеру гістонів. N-кінцеві хвости гістонів Н3 та Н2В виходять за межі нуклеосоми через канали, сформовані маленькими жолобками двох дуплексів сусідніх витків нуклеосомної суперспіралі (рис. 10). Ділянки хвостів, безпосередньо розташованих у каналах, є позитивно зарядженими і, таким чином, додатково скріплюють сусідні витки суперспіралі. N-кінцеві хвости гістонів Н4 і Н2А також взаємодіють з маленьким жолобком зовні нуклеосомної суперспіралі.
Особливе місце –– входу / виходу нуклеосомної ДНК (рис. 10) займає найдовший N-кінцевий хвіст гістону Н3. Позитивно заряджений хвіст Н3 стабілізує структуру нуклеосоми в цій зоні, де спостерігається найвища щільність негативних зарядів ДНК. Значна частина гістонових хвостів просто виходить за межі нуклеосоми. Завдяки своїй структурній лабільності вони беруть участь в організації хроматину на наднуклеосомному рівні, а також відіграють важливу роль платформи для зв’язування різноманітних білків, що залежить від посттрансляційних модифікацій хвостів (див. нижче).
Аналіз структури нуклеосоми та молекулярних механізмів її стабілізації дозволяє сформулювати такі положення:
• Електростатичні взаємодії між ДНК і гістонами за фізіологічних умов є дуже міцними – остаточне руйнування нуклеосоми in vitro відбувається при концентрації солі 2 моль/л. Тобто за фізіологічних умов зв’язування гістонів з ДНК є необоротним (неможлива рівновага між зв’язаними та дисоційованими гістонами). Хоча структура нуклеосоми відповідає мінімуму вільної енергії, цього мінімуму неможливо досягти за розумний проміжок часу: гістони швидко й безладно зв’язуються з ДНК без подальшої дисоціації. Для реалізації рівноважних умов ДНК-гістонової взаємодії in vivo у хроматині існують проміжні акцептори гістонів, що виконують роль факторів збирання / руйнування нуклеосом. Ці акцептори мають спорідненість до гістонів дещо меншу, ніж спорідненість гістонів до ДНК (рис. 11), що й забезпечує можливість рівноважного обміну гістонами між ДНК і проміжними акцепторами із зсувом цієї рівноваги в бік ДНК-гістонових комплексів.
• Незважаючи на міцність ДНК-гістонових взаємодій, тимчасове порушення їх на кінцях нуклеосомної ДНК є можливим за фізіологічних умов: з одного боку, взаємодії на кінцевих ділянках є більш слабкими, ніж у центральній частині нуклеосомної ДНК; з іншого – висока густина негативного заряду (контакт між сусідніми витками нуклеосомної суперспіралі) дестабілізує нуклеосому ДНК на виході з нуклеосоми. Це приводить до тимчасового часткового розкручування нуклеосомної ДНК на кінцях, що є одним із головних шляхів структурної динаміки нуклеосом.
• Наявність проміжних акцепторів гістонів робить можливим обмін димерами Н2АН2В між різними нуклеосомами: тимчасове видалення димерів є іншим важливим шляхом структурної динаміки хроматину.
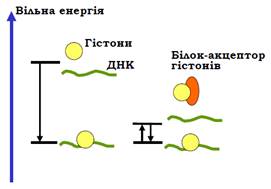
Рис. 11. Роль проміжних акцепторів гістонів у збиранні нуклеосоми: дисоціація гістонів є практично забороненою за фізіологічної іонної сили; у присутності акцептора взаємодія гістонів з ДНК стає рівноважною.
• Незважаючи на те, що ДНК-гістонові взаємодії в нуклеосомі є неспецифічними щодо послідовності пар основ ДНК (електростатичні взаємодії між фосфатами ДНК і позитивними амінокислотними залишками), у хроматині спостерігається феномен переважного позиціювання нуклеосом відносно послідовності. Основною причиною позиціювання нуклеосом є залежність від послідовності здатності ДНК до деформацій, які супроводжують формування нуклеосоми. У результаті нуклеосома обирає таку ділянку, де вигин ДНК потребує менших енергетичних витрат. Це приводить до важливих функціональних наслідків – диференційного експонування ділянок ДНК до дії регуляторних факторів.
• При функціонуванні хроматину виникає необхідність у змінах згаданого вище експонування / екранування певних ділянок ДНК – репозиціюванні нуклеосом. Міцність ДНК-гістонових взаємодій не дозволяє спонтанного зсуву нуклеосом уздовж ДНК, що зумовлює потребу в особливих АТР-залежних молекулярних пристроях, які здійснюють таке репозиціювання, – факторах ремоделювання хроматину, що також можуть виступати факторами збирання / руйнування нуклеосом.
Дата добавления: 2015-09-11; просмотров: 3624;