Синтез нуклеиновых кислот и белка
При рассмотрении митоза и мейоза мы говорили, что в интерфазе происходит удвоение ДНК. Теперь же посмотрим, как происходит этот процесс. Синтез ДНК (репликация, т.е. копирование) осуществляется в присутствии трифосфатов всех четырех дезоксинуклеотидов, ДНК-затравки и ДНК-полимеразы. Репликация осуществляется полуконсервативным способом, который заключается в том, что молекула ДНК удваивается путем репликации каждой из двух ее цепей. Поэтому каждая двойная спираль ДНК состоит из старой и новой полинуклеотидных цепей.
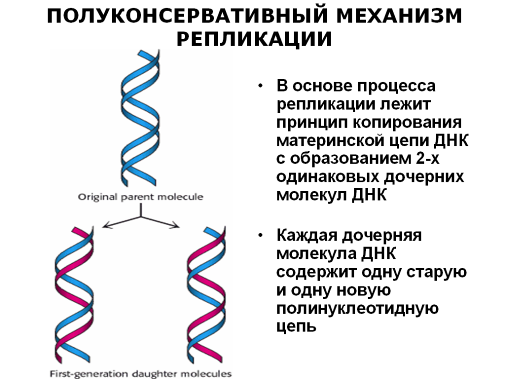
В процессе репликации двуспиральная ДНК локально расплетается в нескольких местах одновременно. В этих местах начинается репликация, которая продолжается до тех пор, пока не достигнет начала другой репликационной вилки. Новая цепь синтезируется ДНК-полимеразой и на всех участках соблюдается полярность сборки полимера: считка идет от 3’-конца одной цепи к ее 5’-концу, а комплементарная цепь синтезируется в направлении от 5’- к 3’-концу.
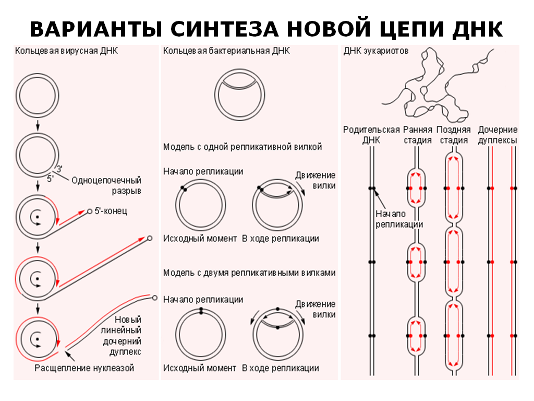
В процессе репликации ДНК взаимодействует несколько десятков ферментов и различных факторов. Следует обратить внимание на точность, с которой происходит процесс репликации. В среднем в процессе репликации одна ошибка приходится на 3 миллиарда пар оснований при скорости репликации от 500 нуклеотидов в секунду у бактерий до 50 нуклеотидов у млекопитающих.

Теперь рассмотрим вопрос о том, как же хранящаяся в ДНК информация трансформируется в структуру белка. Структура любой белковой молекулы закодирована в ДНК, которая непосредственного участия в ее синтезе не принимает. Процесс биосинтеза белка осуществляется на рибосомах, расположенных преимущественно в цитоплазме. Следовательно, для передачи генетической информации с ДНК к месту синтеза белка требуется посредник. Таким посредником и является иРНК.
Синтез РНК-копии нуклеотидной последовательности того или иного участка ДНК катализируется ферментом, называемым РНК-полимераза. А сам процесс синтеза носит название транскрипция. Транскрипция осуществляется в ядре и начинается с присоединения РНК-полимеразы к специфической последовательности ДНК, к так называемому промотору, отмечающему то место на ДНК, с которого должен начаться синтез РНК.

Присоединившись к промотору, фермент раскручивает короткий участок ДНК, соединяет между собой рибонуклеозидтрифосфаты. Продвигаясь вдоль матричной цепи ДНК, фермент постепенно наращивает комплементарную цепь иРНК в направлении от 5’- к 3’-концу, добавляя к ней последовательно по одному нуклеотиду.
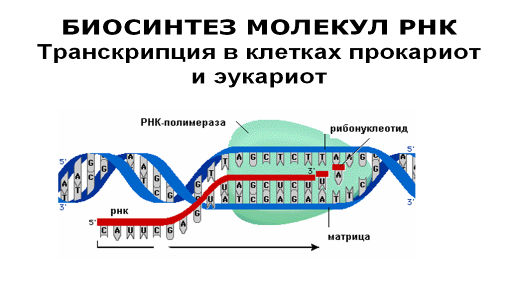
Фермент продолжает присоединять нуклеотиды к растущей цепи до тех пор, пока не встретит на своем пути еще одну специфическую нуклеотидную последовательность в цепи ДНК, так называемый стоп-сигнал, или сигнал терминации транскрипции. Достигнув этой точки, РНК-полимераза отделяется и от матричной ДНК, и от новосинтезированной РНК.
В принципе любой участок ДНК может быть транскрибирован с образованием двух различных молекул РНК – по одной на каждую из двух цепей двойной спирали ДНК. В действительности же транскрибируется только одна из двух цепей ДНК, хотя не всегда одна и та же для соседних генов. Поскольку цепь ДНК, выполняющая роль матрицы, должна считываться от 3’-конца к 5’-концу, выбор одной из двух цепей ДНК в качестве матрицы для синтеза РНК определяется направлением движения РНК-полимеразы. В свою очередь направление движения РНК-полимеразы задается ориентацией нуклеотидной последовательности промотора, с которого РНК-полимераз начинает считывание. Обычно в любом участке двойной спирали ДНК транскрибируется только одна из двух цепей ДНК, хотя бывают и случаи, когда обе цепи несут информацию о структуре белков. Отметим, что каждая синтезированная молекула РНК идентична по своей полярности и нуклеотидной последовательности (если не считать замены Т на У) не той цепи, которая служит для нее матрицей, а второй, которая комплементарна первой.
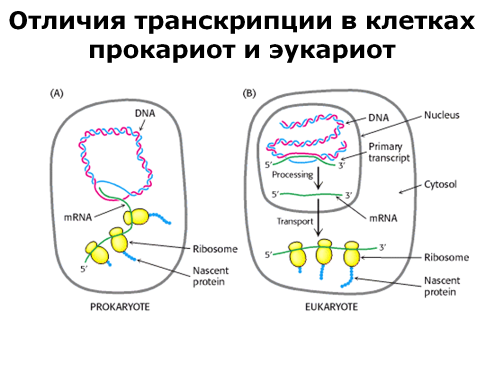
Только что рассмотренный механизм транскрипции характерен для прокариот, и хотя транскрипция у эукариот в принципе не отличается от таковой у прокариот, однако он содержит дополнительные стадии, усложняющие его. Во-первых, у эукариот существует три вида РНК-полимераз, каждая из которых принимает участие в синтезе определенного вида РНК. Гены, кодирующие белки, транскрибируются одним видом РНК-полимеразы. А гены, которые кодируют РНК, имеющие самостоятельное функциональное значение (рРНК и тРНК), транскрибируются другими видами РНК-полимеразы; обычно эти гены представлены в геноме большим числом копий. Синтез рибосомальной РНК осуществляется в ядрышке, которое также служит местом сборки всех клеточных рибосом.
Во-вторых, в процессе транскрипции происходит одновременная модификация получаемой нити РНК. В-третьих, как мы уже говорили, ДНК эукариот содержит некодирующие (интронные) последовательности, и которые естественно входят в первичный РНК-транскрипт. Следовательно, перед тем, как с РНК будет считана информация о структуре белка должны быть удалены эти последовательности, что и происходит. Процесс удаления интронов из первичного РНК-транскрипта называется процессингом. В результате этого процесса образуется матричная РНК, – мРНК, пригодная для использования в качестве источника информации для синтеза белка. Процессинг одного и того же РНК-транскрипта может осуществляться разными способами, давая тем самым начало нескольким мРНК, кодирующим разные белки.
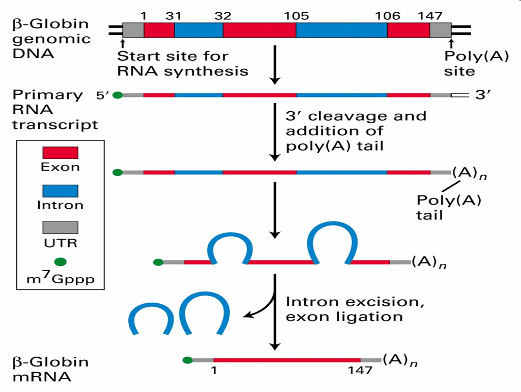
мРНК выходит через поры ядерной оболочки из ядра у месту синтеза белка в цитоплазму. Синтез новых молекул белка осуществляется на рибосомах.

Прежде чем мы рассмотрим процесс трансляции, или синтеза белка, рассмотрим вопрос о том, как информация, записанная четырьмя азотистыми основаниями в мРНК, может быть переведена на белок, информация в котором записана 20 аминокислотами. Таким переводчиком является генетический код. Согласно генетического кода каждая аминокислота в белке кодируется определенным сочетанием трех расположенных рядом нуклеотидов (триплет)матричной РНК, называемых кодоном. Код является вырожденным. Под вырожденностью кода понимают возможность включения в белковую молекулу одной аминокислоты несколькими триплетами. Триплеты не перекрывают друг друга, один триплет не может входить в состав других, каждый из них самостоятельно кодирует аминокислоту. Поэтому в полипептидной цепи рядом могут находится любые две аминокислоты, и возможны какие угодно их сочетания. И, наконец, код не имеет знаков препинания, это означает, что если произойдет выпадение одного нуклеотида, то при считывании его место займет ближайший нуклеотид из соседнего кодона, благодаря чему изменится весь порядок считывания. Поэтому правильное считывание кода с матричной РНК обеспечивается только в том случае, если он считывается со строго определенного кодона.

Нуклеотидный состав универсален для всех живых организмов, что свидетельствует о единстве происхождения всех живых организмов.
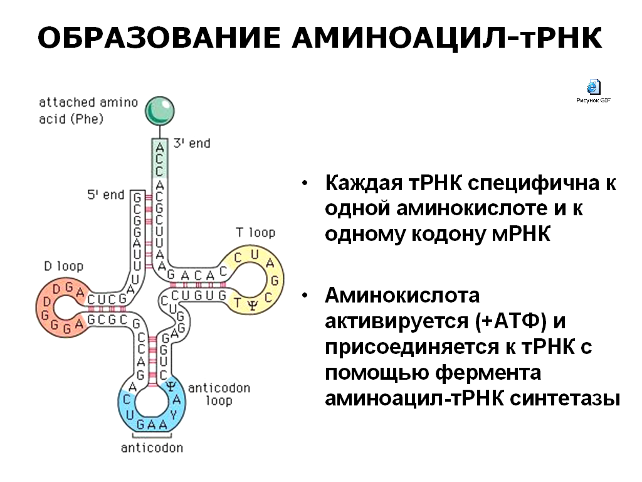
Процесс белкового синтеза протекает в пять основных этапов, каждый из которых требует ряда компонентов, причем в эукариотических и прокариотических клетках имеет место один и тот же механизм с небольшими отличиями в деталях. Первый этап, представляющий собой активацию аминокислот, протекает не в рибосоме, а в цитозоле, где каждая из 20 аминокислот присоединяется к определенной тРНК, представляющей собой сравнительно небольшие одноцепотчатые молекулы, структурная формула которой похожа на форму кленового листа.
Каждая тРНК имеет акцепторный конец, к которому присоединяется активированная аминокислота. Активацию аминокислот осуществляют специфические ферменты аминоацил-тРНК-синтетазы, т.е. для каждой аминокислоты существует свой фермент. Механизм активации заключается в том, что фермент одновременно взаимодействует с соответствующей аминокислотой и с АТФ, которая при этом теряет пирофосфат. Тройной комплекс из фермента, аминокислоты и АТФ, называемый активной аминокислотой, способен спонтанно образовывать пептидную связь. Этот процесс активации – необходимый этап белкового синтеза, поскольку свободные аминокислоты не могут прямо присоединяться к полипептидной цепи. В противоположной части молекулы тРНК располагается специфический триплет (антикодон), ответственный за прикрепление по принципу комплементарности к определенному триплету мРНК (кодону). Каждая тРНК имеет свой антикодон, соответствующий транспортируемой аминокислоте.

На втором этапе – инициации полипептидной цепи, мРНК, содержащая информацию о данном полипептиде, связывается с рибосомой, а затем и с инициирующей аминокислотой, прикрепленной к соответствующей тРНК. Эта РНК, несущая инициирующую кислоту, взаимодействует по принципу комплементарности с находящимся в составе мРНК особым триплетом, или кодоном, который сигнализирует о начале полипептидной цепи. На третьем этапе – элонгации, полипептидная цепь удлиняется с N-конца в сторону C-конца за счет последовательного ковалентного присоединения аминокислот, каждая из которых доставляется к рибосоме и встраивается в определенное положение с помощью соответствующей тРНК, образующей комплементарные пары с отвечающим ей кодоном в мРНК.

После завершения синтеза полипептидной цепи, о котором сигнализирует терминирующий кодон мРНК, наступает четвертый этап – терминация и освобождение, в котором происходит высвобождение полипептида из рибосомы.

Освободившийся полипептид вступает в пятый этап, –сворачивая полипептидной цепи и процессинг. Чтобы принять свою нативную биологически активную форму, полипептид должен свернуться, образуя при этом определенную пространственную конфигурацию. До и после сворачивания новосинтезированный полипептид претерпевает процессинг, заключающийся в удалении инициирующих аминокислот, в отщеплении лишних аминокислотных остатков, во введении в определенные аминокислотные остатки различных групп, а также в присоединении олигосахаридов или простетических групп.
Для ускорения синтеза белка часто с одной молекулы мРНК происходит одновременное считывание информации сразу несколькими рибосомами. В зависимости от того, для каких целей синтезируется белок, трансляция проходит либо на свободных рибосомах, либо на рибосомах прикрепленных к эндоплазматическому ретикулуму. В последнем случае синтезируемая полипептидная цепь сразу помещается внутрь ЭПР.

Как можно видеть из всего вышесказанного, процесс синтеза белка имеет сложный механизм. В этом процессе принимает участие до трех сотен различных веществ. На образование пептидной связи затрачивается не менее 29,2 ккал энергии фосфатной группы, в то время как стандартная свободная энергия ее гидролиза составляет всего около 5 ккал. Таким образом, чистая затрата энергии на синтез пептидной связи составляет 24,2 ккал/мол. Столь высокий расход энергии служит одним из важных факторов, обеспечивающих почти совершенную точность биологического перевода генетической информации мРНК на язык аминокислотной последовательности белков.
Дата добавления: 2015-11-16; просмотров: 3510;