Клеточное дыхание и фотосинтез
Вот теперь мы подошли к вопросу о том, откуда и как клетка получает энергию, как ее преобразует. И начнем, пожалуй, с рассмотрения понятия метаболизма.
Совокупность химических реакций, протекающих в организме, называется метаболизмом или обменом вещества
По типу обмена веществ организмы подразделяются на две группы: автотрофы и гетеротрофы.
Автотрофы — организмы, способные синтезировать органические вещества из неорганических и использующие для этого синтеза энергию солнца (фотоавтотрофы) или энергию, выделяющуюся при окислении неорганических веществ (хемоавтотрофы).
Гетеротрофы — организмы, использующие для своей жизнедеятельности органические вещества, синтезированные другими организмами.
Метаболизм представляет собой высоко скоординированную и целенаправленную клеточную активность, обеспечиваемую участием многих взаимосвязанных мультиферментных систем.
Метаболизм в организме выполняет четыре специфические функции:
1) снабжение химической энергией, которая добывается путем расщепления богатых энергией пищевых веществ, поступающих в организм из среды, или путем преобразования улавливаемой солнечной энергии;
2) превращение молекул пищевых веществ в строительные блоки, которые используются в дальнейшем клеткой для построения макромолекул;
3) сборку белков, нуклеиновых кислот, липидов, полисахаридов и прочих клеточных компонентов из этих строительных блоков;
4) синтез и разрушение тех биомолекул, которые необходимы для выполнения каких-либо специфических функций данной клетки.
Хотя метаболизм слагается из сотен различных ферментативных реакций, центральные метаболические пути немногочисленны и почти у всех живых форм в принципе едины. Метаболизм включает катаболические и анаболические пути.
Катаболизм (энергетический обмен, диссимиляция), – это фаза метаболизма, в которой происходит расщепление сложных органических веществ до более простых конечных продуктов.
Реакции катаболизма сопровождаются выделением энергии. Энергия, освобождающаяся при распаде органических веществ, не сразу используется клеткой, а запасается в форме АТФ и других высокоэнергетических соединений.. Синтез АТФ происходит в клетках всех организмов в процессе фосфорилирования — присоединения неорганического фосфата к АДФ.
АТФ — универсальный источник энергообеспечения клетки.
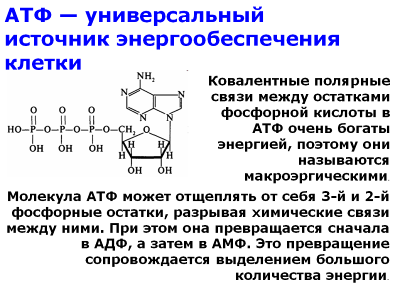
Часть ее запасается также в богатых энергией водородных атомах коферментов: никотинамидадениндинуклеотидфосфата, находящегося в восстановленной форме (НАДФН), никотинамидадениндинуклеотида (НАДН), флавинадениндинуклеотида (ФАДН2).
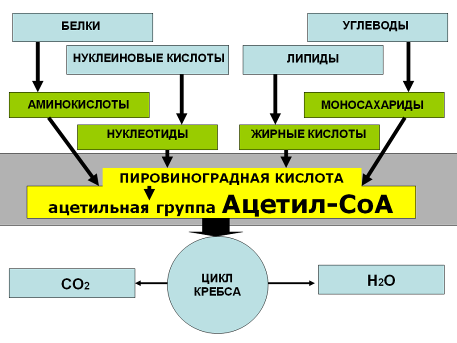
Ферментативное расщепление тех главных питательных веществ, которые служат в клетке источником энергии, совершается постепенно – через ряд последовательных ферментативных реакций, которые можно условно разделить на три стадии. На первой стадии сотни белков и многие виды полисахаридов и липидов расщепляются на составляющие их строительные блоки. На второй стадии эти строительные блоки превращаются в один общий продукт – ацетильную группу ацетил-СоА. Третья стадия – это общий путь всех катаболических путей – цикл лимонной кислоты (цикл Кребса) – образование всего лишь трех конечных продуктов: воды, углекислоты и энергии.
Подготовительный этап.
Заключается в ферментативном расщеплении сложных органических веществ до простых: белковые молекулы — до аминокислот, жиры — до глицерина и карбоновых кислот, углеводы — до глюкозы, нуклеиновые кислоты — до нуклеотидов. Распад высокомолекулярных органических соединений осуществляется или ферментами желудочно-кишечного тракта или ферментами лизосом. Вся высвобождающаяся при этом энергия рассеивается в виде тепла. Образовавшиеся небольшие органические молекулы могут быть использованы в качестве «строительного материала» или могут подвергаться дальнейшему расщеплению.
Бескислородное окисление, или гликолиз.
Этот этап заключается в дальнейшем расщеплении органических веществ, образовавшихся во время подготовительного этапа, происходит в цитоплазме клетки и в присутствии кислорода не нуждается. Главным источником энергии в клетке является глюкоза. Процесс бескислородного неполного расщепления глюкозы — гликолиз.
При гликолизе молекула глюкоза, содержащая шесть атомов углерода, претерпевает ряд превращений, в результате которых распадается на две молекулы пирувата, содержащих по три атома углерода. Для такого превращения требуется десять последовательных ферментативных реакций, в которых происходит образование ряда промежуточных фосфатсодержащих соединений
Последовательность реакций гликолиза можно разделить на два этапа. На первом, подготовительном, этапе (реакции 1–5) происходит фосфорилирование глюкозы и ее расщепление на две фосфотриозы. Поскольку глюкоза стабильное соединение, на ее активацию необходима затрата энергии. Для расщепления одной молекулы глюкозы требуется израсходовать две молекулы АТФ
На втором этап гликолиза, также состоящем из пяти реакций, энергия, высвобождающаяся при превращении двух молекул глицеральдегид-3-фосфата в две молекулы пировиноградной кислоты (пирувата), в результате сопряженного фосфорилирования четырех молекул АДФ запасается в виде четырех молекул АТФ. Кроме того, на втором этапе гликолиза на каждую из двух молекул фосфотриоз восстанавливается по одной молекуле НАДН.
С6Н12О6 + 2АДФ + 2Н3РО4 + 2НАД+ →
2С3Н4О3 + 2АТФ + 2Н2О + 2НАД·Н2.

Аминокислоты, нуклеотиды, моносахариды и карбоновые кислоты, образующиеся на первой стадии катаболизма после предварительной модификации также включаются в гликолиз, выделяя энергию и в конечном счете превращаются в молекулу пировиноградной кислоты
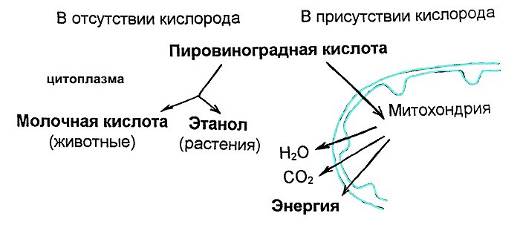
Важная роль пирувата в катаболизме углеводов определяется тем, что это соединение лежит в точке пересечения различных катаболических путей. При аэробных условиях в животных тканях продуктом гликолиза является пируват, а НАДН окисляется за счет молекулярного кислорода, передавая свой атом водорода в электронтранспортную цепь митохондрий, где с его помощью синтезируются три молекулы АТФ. Поскольку при гликолизе одной молекулы глюкозы образуется две молекулы НАДН, то, таким образом, всего в процессе гликолиза (при условии последующего окисления НАДН) образуется восемь молекул АТФ.
Иначе обстоит дело в анаэробных условиях, например, в напряженно работающих скелетных мышцах или в клетках молочнокислых бактерий. В этих условиях образовавшийся при гликолизе НАДН окисляется не за счет кислорода, а за счет пирувата, восстанавливая его в лактат, т.е. молочную кислоту.
У молочнокислых бактерий молочная кислота остается конечным продуктом гликолиза.
У дрожжей и ряда других микроорганизмов, образовавшийся в процессе гликолиза пируват сбраживается до этанола и углекислого газа с одновременным окислением НАДН.
В результате гликолиза одного моля глюкозы высвобождается 200 кДж энергии, из которых 120 кДж рассеивается в виде тепла, а 80 кДж запасается в АТФ
Анаэробное образование АТФ из глюкозы в реакциях гликолиза относительно неэффективно. Конечные продукты анаэробного гликолиза все еще несут в себе очень большое количество химической энергии, которая может быть выделена если эти продукты окислить. Развитие окислительного катаболизма в аэробных микроорганизмах и митохондриях эукариотических клеток стало возможно лишь после того, как в результате фотосинтеза, осуществляемого цианобактериями, в атмосфере Земли было накоплено достаточное количество молекулярного кислорода.
У большинства современных организмов, дышащих кислородом, пируват не превращается в лактат, а утилизируется дальше. Он поступает в каскад ферментативных реакций, в ходе которого расходуется кислород, образуется углекислота и синтезируется АТФ. Все эти реакции в сумме называются клеточным дыханием.
Обратим ваше внимание на то, что клеточное дыхание состоит из двух процессов. В ходе одного из них углерод окисляется до углекислого газа, но молекулярный кислород не расходуется – атомы кислорода берутся из органических веществ и воды, которая здесь не образуется, а расходуется. При этом образуются излишки водорода, которые идут на восстановление коферментов. В ходе второго процесса коферменты окисляются и отдают водород (который сначала разделяется на протоны и электроны, имеющие разную судьбу), именно здесь он связывается с молекулярным кислородом с образованием воды. АТФ образуется преимущественно в ходе второго процесса. Первый процесс называется циклом трикарбоновых кислот, или циклом Кребса, второй – окислительным фосфорилированием.
Рассмотренный выше процесс гликолиза происходит в цитоплазме. Клеточное же дыхание происходит в митохондриях. Для этого продукт гликолиза – пируват - должен попасть внутрь митохондрий.
Итак, мы в митохондрии. Каскад реакций клеточного дыхания начинается с реакции, одним из субстратов которой является пируват, а одним из продуктов – ацетилкофермент-А, или ацетил-coA. Ацетил-coA – одно из важнейших веществ в биохимических путях. Он образуется в процессе расщепления сахаров, жирных кислот и некоторых аминокислот и используется при их синтезе. Во всех этих случаях он является реакционноспособным носителем ацетильной группы. В одних реакциях она используется для синтеза органических веществ, в других – для их «сжигания» в качестве топлива. Поэтому ацетил-coA является важнейшим посредником во множестве биохимических процессов, связанных с обменом вещества и энергии. Посмотрим на это замечательное вещество.
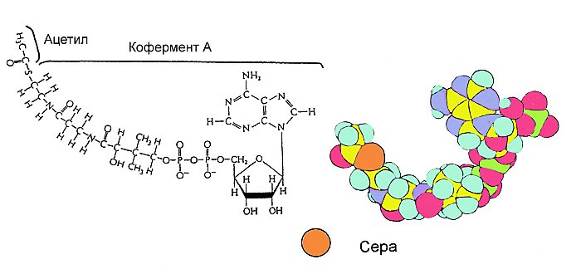
Мы снова видим знакомый нуклеотид аденозин, потом – довольно длинную углеводородную цепь, включающую атомы азота и заканчивающуюся атомом серы, к которой и присоединена ацетильная группа. (Молекула без ацетильной группы – это просто кофермент А.)
Ацетил-coA образуется с затратой молекулы пирувата в ходе сложной реакции, катализируемой целым комплексом из трех ферментов и пяти коферментов, прикрепленным к митохондриальной мембране – пируватдегидрогеназным комплексом. При этом от молекулы пирувата отщепляется молекула диоксида углерода, а оставшаяся от него ацетильная группа присоединяется к коферменту А, с образованием ацетил-соА. Реакция имеет энергетический выигрыш, который идет на восстановление одной молекулы НАД+ до НАД-H. В этой реакции мы впервые видим, как атом углерода переходит из органического вещества в углекислый газ.
Ацетил Co-A вступает в циклический биохимический процесс, называющийся циклом Кребса. Он назван так по имени Ганса Кребса, описавшего его в 1937 г., за что он впоследствии получил Нобелевскую премию.
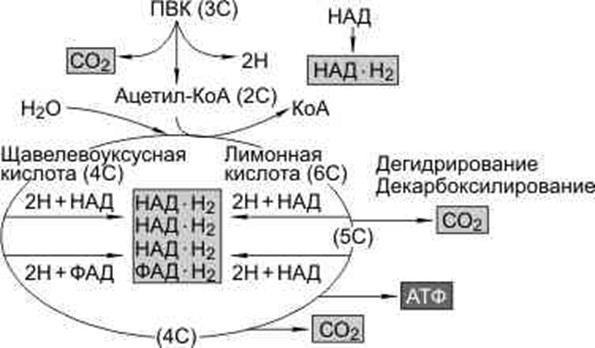
Цикл представляет собой 10 последовательных химических реакций, в ходе которых 10 органических кислот последовательно преобразуются одна в другую. В одном месте в этот цикл входит уже знакомый нам ацетил-соА, который отдает свою ацетильную группу оксалоацетету (щавелевоуксусной кислоте), в результате чего образуется цитрат (лимонная кислота). Если первая молекула содержала четыре атома углерода, то вторая соответственно содержит уже шесть (в ацетильной группе имеется два углерода). Цикл Кребса замыкается, когда в конце концов мы приходим к тому же оксалоацетату с его четырьмя атомами углерода, к которому может снова присоединяется ацетильная группа от ацетил-соА.
В ходе последовательных превращений всех этих кислот происходят события нескольких типов:
– кислоты теряют два атома углерода за счет образования двух молекул углекислого газа;
– кислоты присоединяют две молекулы воды;
– излишки водорода уходят на восстановление трех молекул НАД+ до НАД-H, а также на восстановление еще одного кофермента – флавинадениндинуклеотида (ФАД) до ФАД-Н2;
Суммарная реакция гликолиза и разрушения ПВК в митохондриях до водорода и углекислого газа выглядит следующим образом:
С6Н12О6 + 6Н2О → 6СО2 + 4АТФ + 12Н2
Две молекулы АТФ образуются в результате гликолиза, две — в цикле Кребса; две пары атомов водорода (2НАДЧН2) образовались в результате гликолиза, десять пар — в цикле Кребса.
Все вещества цикла Кребса – и кислоты, и катализирующие реакции ферменты – находятся в одном и том же растворе внутри митохондрий, поэтому цикл не имеет пространственного содержания – это просто последовательность превращений веществ. Он играет центральную роль в метаболизме клетки, так как участвующие в нем вещества являются промежуточными веществами многих метаболических процессов. Этот цикл задействован в расщеплении и синтезе углеводов, в расщеплении и синтезе жирных кислот, в расщеплении и синтезе многих аминокислот, в синтезе азотистых оснований нуклеотидов и других важных веществ.
Слайд Последним этапом является окисление пар атомов водорода с участием кислорода до воды в цепи переноса электронов (ЦПЭ) с одновременным фосфорилированием АДФ до АТФ.
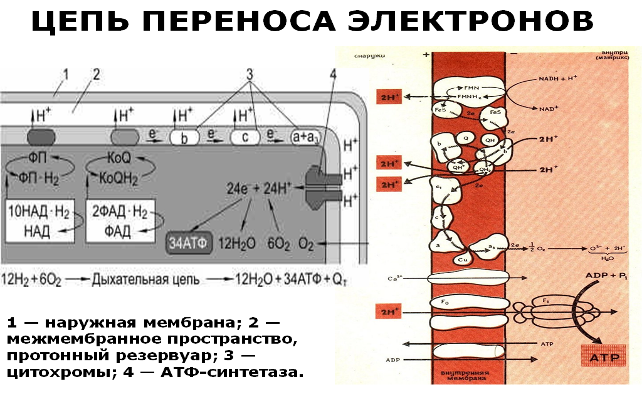
Водород передается трем большим ферментным комплексам (флавопротеины, коферменты Q, цитохромы) дыхательной цепи, расположенным во внутренней мембране митохондрий. У водорода отбираются электроны, которые в матриксе митохондрий в конечном итоге соединяются с кислородом:
О2 + e— → О2—.
Протоны закачиваются в межмембранное пространство митохондрий, в «протонный резервуар». Внутренняя мембрана непроницаема для ионов водорода, с одной стороны она заряжается отрицательно (за счет О2—), с другой — положительно (за счет Н+). Когда разность потенциалов на внутренней мембране достигает 200 мВ, протоны проходят через канал фермента АТФ-синтетазы, образуется АТФ, а цитохромоксидаза катализирует восстановление кислорода до воды. Так в результате окисления двенадцати пар атомов водорода образуется 34 молекулы АТФ.
АТФ образуется в митохондриях, но нужна всей клетке. Однако образовавшаяся АТФ не может самопроизвольно проникать из митохондрий в цитоплазму. Для этого в митохондриальной мембране имеется специальным белок – транслоказа, который производит реакцию обмена одной молекулы АТФ изнутри митохондрии на одну молекулу АДФ снаружи митохондрии, причем делает это безвоздмездно, то есть без затрат энергии.
Суммарная реакция расщепления глюкозы до углекислого газа и воды выглядит следующим образом:
С6Н12О6 + 6О2 → 6СО2 + 6Н2О + 36АТФ + Qт,
где Qт — тепловая энергия.
Итак, если учесть все реакции, предшествующие образованию ацетил-СоА, окажется, что полное окисление одной молекулы глюкозы дает 36 молекул АТФ. Это максимальное значение, так как фактически количество синтезируемой АТФ зависит от того, какая доля энергии протонного градиента идет на синтез АТФ, а не на другие процессы. Если сравнить изменение свободной энергии при прямом сгорании углеводов с общим количеством энергии, запасаемой в АТФ, то окажется, что эффективность преобразования энергии питательных веществ в энергию АТФ превышает 50%, что значительно больше эффективности большинства энергопреобразующих устройств, созданных человеком.
Если вернуться к внутренней мембране митохондрии, то можно видеть, что обратный перенос протона через мембрану теоретически возможен и без сопряжения реакции фосфорилирования. Это явление существует на самом деле. В таких случаях вся энергия, поступившая в цепь переноса электронов рассеивается в виде тепла. Возможность такого способа получения тепла используется организмами.
У многих млекопитающих, в том числе и у человека, имеется особый тип жировой ткани, называемый бурым жиром. Цвет этой ткани обусловлен большим содержанием митохондрий. Митохондрии бурого жира отличаются от всех прочих митохондрий этого организма тем, что содержат в мембране специальный белок, который проводит электрон. Через этот белок идет утечка электронов в межмембранное пространство. В итоге наблюдается “холостая” циркуляция протонов и вместо АТФ происходит выделение тепла.
Бурый жир составляет менее 1-2% массы тела. Тем не менее стимуляция этой ткани при охлаждении животного повышает ее теплопродукцию до 400 вт на кг веса, что намного больше обычной теплогенной способности тканей млекопитающих (человек в состоянии покоя образует около 1 вт тепла на кг массы). Если тепла не надо, то этот белок закрывает канал и митохондрия синтезирует АТФ.
В организме человека бурый жир сосредоточен в верхней части спины ближе к шее. Он окружает кровеносные сосуды, питающие мозг кровью, поэтому теплопродукция в буром жире имеет огромное значение для выживания организма на холоде.
Итак, начальные этапы окисления глюкозы (гликолиз) начинается в цитозоле, а заканчивается окисление в митохондрии. Митохондрия служит одновременно и силовой станцией клетки, и местом, где происходит окончательное окисление атомов углерода и водорода молекул питательных веществ. Митохондрия – это центр, к которому ведут все катаболические пути независимо от того, что служит для них первоначальным субстратом – сахара, жиры или белки. Объясняется это тем, что не только пируват, но и жирные кислоты, равно как и некоторые аминокислоты, тоже поступают из цитозоля в митохондрии, где они превращаются в ацетил-СоА или в один из промежуточных продуктов цикла лимонной кислоты.
Помимо образования АТФ, необходимой для процессов биосинтеза, митохондрия служит еще и отправной точкой биосинтетических реакций, поскольку промежуточные продукты цикла лимонной кислоты являются исходными продуктами для синтеза многих жизненно важных веществ.
Анаболизм, - это та фаза метаболизма, в которой из малых молекул синтезируются большие. Анаболизм, как и катаболизм, протекает в те же три стадии, но в обратном порядке. Поскольку биосинтез – это процесс, в результате которого увеличиваются размеры молекул и усложняется их структура, он требует затраты свободной энергии. Источником этой энергии служит распад АТФ до АДФ и неорганического фосфата. Для биосинтеза некоторых клеточных компонентов требуются также богатые энергией водородные атомы, донором которых является НАДФН.
Катаболические и анаболические реакции протекают в клетках одновременно, но катаболический путь и соответствующий ему, но противоположный по направлению анаболический путь между данным предшественником и данным продуктом обычно не совпадают. Причин для этого, по крайней мере, две. Первая из них заключается в том, что путь, по которому идет расщепление той или иной биомолекулы, может быть по энергетическим соображением непригодным для ее биосинтеза. Вторая причина состоит в том, что эти последовательности реакций должны регулироваться раздельно, следствием чего является необходимость быть различными хотя бы на одной из ферментативных стадий. Таким образом, хотя соответствующие катаболические и анаболические пути неидентичны, однако их связывает общая стадия (цикл лимонной кислоты), называемая амфиболической стадией метаболизма, поскольку она выполняет двойную функцию. В катаболизме на этой стадии завершается распад сравнительно небольших молекул, а в анаболизме ее роль заключается в поставке небольших молекул-предшественников для биосинтеза.
Связующим звеном между катаболизмом и анаболизмом является не только амфиболическая стадия метаболизма, но и энергия высокоэнергичной фосфодиэфирной связи в АТФ, которая является универсальной системой клетки, служащей для переноса энергии, а также богатый энергией атом водорода в коферментах.
Клеточный метаболизм основан на принципе максимальной экономии. Общая скорость катаболизма, обеспечивающего клетку энергией, определяется не просто наличием или концентрацией клеточного топлива; она обусловлена потребностью клетки в энергии в форме АТФ и НАДФН. Клетка потребляет в каждый данный момент как раз такое количество питательных веществ, которое позволяет ей удовлетворять свои энергетические нужды, что в свою очередь, подразумевает наличие очень хорошего механизма регуляции метаболических реакций.
В регуляции метаболических путей участвуют механизмы трех типов. Первый из них, наиболее быстро реагирующий на изменение ситуации, связан с действием аллостерических ферментов, когда продукт последней реакции влияет на активность фермента в начале цепи. Иногда в качестве такого продукта выступает АТФ.
Второй тип механизмов, регулирующих метаболизм у высших организмов, - это гормональная регуляция. Гормонами называют особые химические вещества, вырабатываемые различными эндокринными железами и выделяемые непосредственно в кровь; они переносятся кровью к другим тканям или органам и здесь стимулируют или тормозят определенные виды метаболической активности. Например, в клетках мозгового слоя надпочечников синтезируется и накапливается гормон адреналин. В момент возникновения опасности, вызывающей у животного состояние тревоги и готовности к борьбе или бегству, головной мозг посылает нервный импульс, достигающих мозгового слоя надпочечников, его клетки выделяют адреналин, который попадает в кровь. Концентрация адреналина в крови возрастает за время, исчисляемое секундами, почти в 1000 раз. Адреналин присоединяется к особым рецепторным участкам на поверхности мышечных клеток и клеток печени. Связывание адреналина служит сигналом; этот сигнал передается во внутренние отделы клетки и вызывает здесь модификацию гликоген-фосфорилазы (первого фермента в системе, катализирующей превращение гликогена в глюкозу). Фермент переходит из менее активной формы в более активную, что стимулирует распад гликогена в скелетных мышцах; этот процесс приводит к образованию лактата и к запасанию энергии в форме АТФ. Одновременно адреналин тормозит синтез гликогена в печени из глюкозы, что способствует максимальному поступлению глюкозы в кровь. Кроме того, адреналин ускоряет ритм сердца, увеличивает сердечный выброс и повышает кровяное давление, подготавливая тем самым сердечно-сосудистую систему к активности в экстремальной ситуации. Таким образом, регулируя активность соответствующих ферментов, адреналин приводит животное в состояние готовности к борьбе или к бегству.
Третий тип механизмов, регулирующих метаболизм, связан с изменением концентрации данного фермента в клетке. Концентрация всякого фермента в любой данный момент определяется соотношением скоростей его синтеза и распада. Скорость синтеза некоторых ферментов при определенных условиях резко возрастает; соответственно увеличивается и его концентрация. Если, например, животное получает рацион, богатый углеводами, но бедный белками, то в печени у него оказывается крайне низким содержание ферментов, катализирующих распад аминокислот до ацетил-СоА. Поскольку при таком рационе эти ферменты практически не нужны, они и не вырабатываются в больших количествах. Однако стоит перевести животных на рацион, богатый белком, как уже через сутки заметно повышается содержание этих ферментов. Следовательно, клетки печени обладают способностью включать или выключать биосинтез специфичных ферментов в зависимости от природы поступающих в них питательных веществ.
Теперь мы обратимся к процессу, который является в конечном счете источником почти всей биологической энергии, т.е. к процессу улавливания солнечной энергии фотосинтезирующими организмами и превращению ее в энергию биомассы. На самом деле есть и другие, менее распространенные варианты биосинтеза органики. Однако главный – именно фотосинтез, в результате которого на Земле ежегодно образуется 150 млрд т сахаров.
Еще в начале исследований фотосинтеза было показано, что имеет место группа реакций, зависящих от освещенности и не зависящих от температуры, а есть группа реакций, которая, наоборот, не зависит от освещенности и зависит от температуры. Первая получила название световой стадии фотосинтеза, вторая – темновой стадии фотосинтеза. Не следует понимать это в том смысле, что одна идет днем, а другая – ночью. Оба набора реакций протекают одновременно, просто для одной свет нужен, а для другой – нет.
Для ознакомления со световой фазой фотосинтеза нам необходимо рассмотреть такое химическое явление, как пигменты. Что такое пигменты? Это окрашенные вещества. А почему некоторые вещества окрашены, в то время как большинство веществ бесцветно? Что означает наше видение некоего цвета? Это означает, что от вещества нам приходит свет, в котором соотношение фотонов с разной длиной волны отличается от дневного белого света. Как вы знаете, белый свет представляет собой смесь фотонов в буквальном смысле всех цветов радуги. Окраска света означает преобладание определенных длин волн над другими. Мы рассматриваем вещества при дневном свете. Соответственно, если мы видим вещество окрашенным, значит, оно избирательно поглощает фотоны с определенными длинами волн. Не имея массы покоя, поглощенные фотоны перестают существовать. Куда же девается их энергия? Она идет на возбуждение молекулы, на перевод ее в новое, более энергетически насыщенное состояние.
Чтобы иметь способность поглощать свет и переходить в энергетически насыщенное состояние, молекула должна представлять собой систему, в которой такое состояние возможно. Большая часть органических пигментов представляет собой вещества с регулярным чередованием двойных и одинарных связей между углеродами, т. е. с сопряженными двойными связями. Эти связи образуют резонансные системы, в которых электроны, участвующие в образовании двойных связей (образованных орбиталями, не задействованными в sp2-гибридизации), могут перемещаться по всей системе и находиться в нескольких энергетических состояниях. Число таких состояний и энергия, необходимая для перехода электрона из одного в другое, строго фиксированы для каждой молекулы.
Энергия, различающая состояния электрона в резонансных системах, такова, что близко соответствует энергии фотонов той или иной длины волны в пределах видимой части спектра. Поэтому резонансные системы будут поглощать те фотоны, энергия которых равна или немного больше перевода их электронов в одно из более энергетически насыщенных состояний.
Давайте посмотрим на молекулы некоторых важных для нашего случая пигментов. Для начала на самый важный пигмент – хлорофилл.
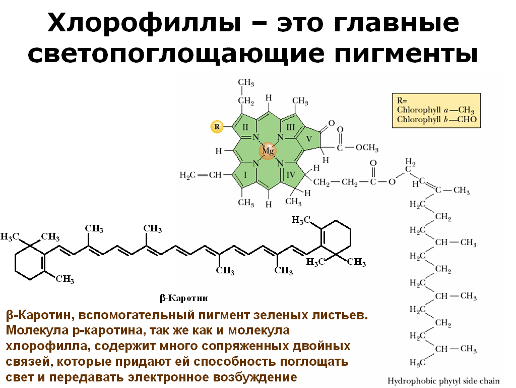
Мы видим ажурную и почти симметричную органическую конструкцию, включающую несколько двойных связей – порфириновое кольцо. В ее центре также находится атом металла, магния. Он связан с четырьмя атомами азота (магний и порфириновое кольцо образуют комплекс). К порфириновому кольцу в молекуле хлорофилла приделан длинный углеводородный хвост. Не имея электроотрицательных атомов, эта часть молекулы неполярна и, следовательно, гидрофобна. При помощи нее хлорофилл заякоривается в гидрофобной средней части фосфолипидной мембраны.
Хлорофилл растений представлен двумя формами – a и b. В зеленых растениях примерно четверть хлорофилла представлена второй формой b. Он отличается тем, что одна метильная группа по краю порфиринового кольца -CH3 замещена на группу -CH2OH. Этого оказывается достаточно, чтобы сместить спектр поглощения молекулы. Эти спектры показаны на рисунке.
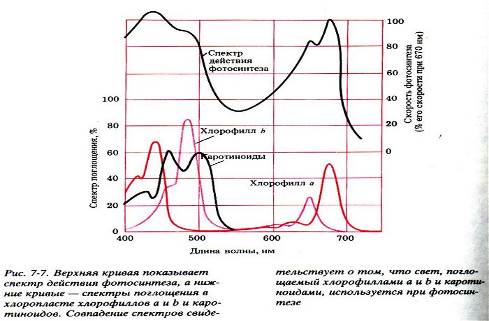
Эта молекула поглощает фотоны в фиолетовой и синей, а затем в красной части спектра, и не взаимодействует с фотонами в зеленой и желтой части спектра. Поэтому хлорофилл и растения выглядят зелеными – они попросту никак не могут воспользоваться зелеными лучами и оставляют их гулять по белу свету (делая его тем самым зеленее). Несколько другую структуру имеют каротиноиды – красные и желтые пигменты. Каротиноиды тоже участвуют в фотосинтезе, но в качестве вспомогательных молекул.
Пигменты фотосинтеза располагаются на внутренней стороне мембраны тилакоидов. Они организованы в фотосистемы – целые антенные поля по улавливанию света – каждая система содержит 250–400 молекул разных пигментов. Но среди них принципиальное значение имеет одна молекула хлорофилла а – она называется реакционным центром фотосистемы. Все остальные молекулы пигментов называются антенными молекулами. Все пигменты в фотосистеме способны передавать друг другу энергию возбужденного состояния.
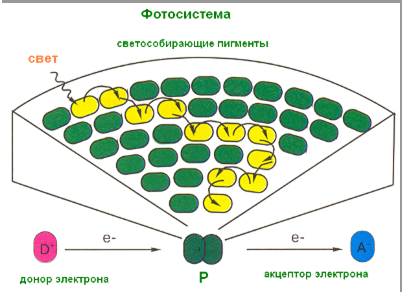
В хлоропластах высших растений содержатся фотосистемы двух типов, каждая со своим набором светособирающих молекул и со своим реакционным центром. Молекулы их реакционных центров несколько различаются – первая имеет максимум поглощения света на длине волны 700 нм, вторая – 680 нм (оговорка сделана для того, чтобы пояснить изображения на схемах), обозначаются они Р700 и Р680. Пространственно эти две фотосистемы расположены в тилакоидной мембране рядом и представляют единое целое.
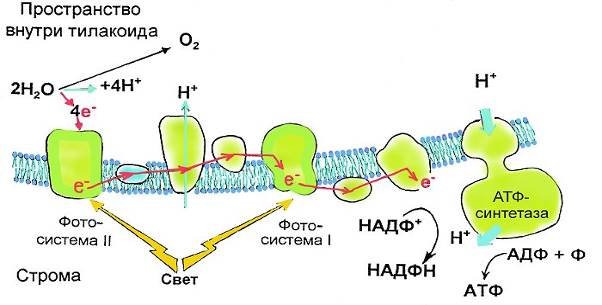
Обычно эти две системы работают сопряженно, как конвейер, состоящий из двух частей и называющийся нециклическим фотофосфорилированием.
Производственный цикл начинается с фотосистемы 2. С ней происходит следующее:
1) антенные молекулы улавливают фотон и передают возбуждение молекуле активного центра Р680;
2) возбужденная молекула Р680 отдает два электрона кофактору Q (очень похожий на тот, который участвует в цепи переноса электронов в митохондриях), при этом она окисляется и приобретает положительный заряд;
3) под действием определенных ферментов, содержащих марганец, окисленная молекула Р680 восстанавливается, отнимая два электрона от молекулы воды. При этом вода диссоциирует на протоны и молекулярный кислород. Для создания одной молекулы кислорода нужно восстановить две молекулы Р680, потерявшие в сумме четыре электрона, при этом образуются четыре протона.
Обратим внимание – именно здесь в ходе фотосинтеза образуется кислород. Поскольку он образуется путем расщепления молекул воды под действием света, этот процесс называется фотолизом воды;
4) эти протоны образуются во внутреннем пространстве тилакоида, где создается избыточная концентрация протонов по сравнению с окружающим пространством (т. е. более кислая среда). Таким образом, формируются наши старые знакомые – протонный градиент и мембранный потенциал. Мы уже знаем, как все это будет использовано:
5) два электрона, поступившие к кофактору Q, передаются далее по цепи переноса электронов через ряд белков. При этом имеет место транспорт протонов против градиента концентрации сквозь мембрану тилакоидов.
Накопленные протоны АТФ-синтетаза будет парами выпускать наружу и синтезировать АТФ из АДФ.
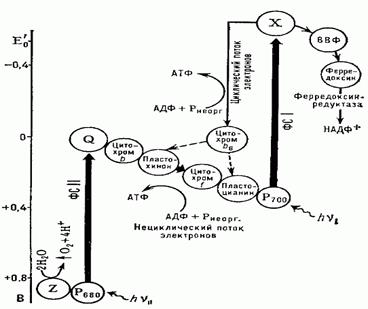
В фотосистеме 1 происходит следующее:
1) антенные молекулы ловят фотон и передают энергию в резонансную систему реакционного центра Р700, который возбуждается и отдает два электрона акцепторному железосодержащему белку (Р430). Как и в случае фотосистемы 2, Р700 тем самым окисляется и приобретает положительный заряд;
2) эта молекула восстанавливается и теряет заряд, получив два «успокоившихся» (но не до исходного состояния – их энергия еще не до конца израсходована!) электрона, изначально поступивших от фотосистемы 2. В этом случае необходимости в фотолизе нет и его не происходит;
3) Р430 отдает электроны другому железосодержащему белку, который называется ферродоксин;
4) получив электроны, этот белок восстанавливает кофермент НАДФ+ до НАДФ-Н. Данный кофермент представляет собой фосфорилированный НАД. Процесс происходит на внешней мембране тилакоида. Для него необходим протон, который берется из внутреннего пространства хлоропласта, внешнему по отношению к тилакоиду. Тем самым протонный градиент только усиливается.
На схеме более или менее показаны все основные процессы световой стадии фотосинтеза.
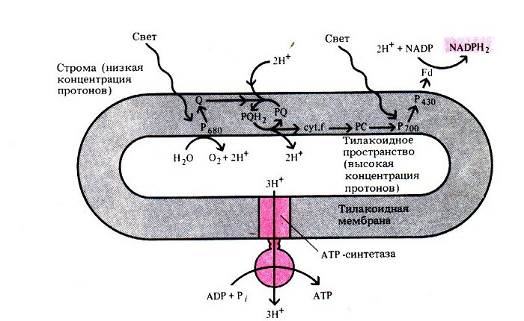
Однако фотосистема 1 может работать и автономно. При этом используется обходной путь переноса электронов от возбужденного реакционного центра – а именно та же цепь переноса электронов, которая ведет из фотосистемы 2. Электроны проходят по ней, вызывая сопряженный транспорт протонов из внешней среды тилакоида во внутреннюю, чем усиливается протонный градиент, и возвращаются обратно к реакционному центру фотосистемы 1 – Р700. Таким образом, здесь свет словно крутит колесо протонного насоса, не окисляя воды и не восстанавливая НАДФ. Это называется циклическим фотофосфорилированием . Оно может идти параллельно с нециклическим. Кроме того, оно используется некоторыми фотосинтетическими бактериями, которые в процессе фотосинтеза не выделяют кислорода.
Результат световой фазы фотосинтеза при нециклическом фотофосфорилировании (а это основной вариант) можно записать в виде такой реакции:
2НАДФ+ 2АДФ + 2Ф- + 2H2O + 4 hv = 2НАДФ-Н + 2АТФ + O2.
Здесь hv - условное обозначение энергии одного фотона, Ф - условное обозначение остатка фосфорной кислоты из раствора.
Итак, мы рассмотрели, откуда в ходе фотосинтеза берется энергия (т. е. АТФ). Осталось рассмотреть, как с использованием этой энергии делается органика.
Растения используют три варианта такого производства. Рассмотрим самый распространенный из них, который к тому же используется сине-зелеными водорослями и фотосинтетическими и даже хемосинтетическими бактериями – цикл Кальвина. Это еще один замкнутый цикл взаимопревращения органических веществ одно в другое под действием специальных ферментов, подобный циклу Кребса. И кстати, еще одна Нобелевская премия, 1961 года – открывшему его Мелвину Кальвину.
Цикл начинается с сахара, имеющего цепочку из пяти атомов углерода и несущего две фосфатные группы – рибулозо-1,5-бифосфат (и им же кончается). Процесс начинается, когда специальный фермент – рибулозобифосфаткарбоксилаза – присоединяет к нему молекулу СО2. Образующаяся на короткое время шестиуглеродная молекула немедленно распадается на две молекулы глицерат-3-фосфата (он же 3-фосфоглицерат, с этим веществом мы уже встречались в гликолизе). Каждая из них содержит три атома углерода (поэтому цикл Кальвина называется также C3-путь фиксации углекислого газа).

Фактически фиксацию угелкислого газа в органике осуществляет именно этот фермент – рибулозобифосфаткарбоксилаза. Это на удивление медленный фермент – он карбоксилирует всего три молекулы рибулозо-1,5-бифосфата в секунду. Для фермента это очень мало! Поэтому самого данного фермента требуется много. Он фиксирован на поверхности тилакоидных мембран и составляет около 50 % от всех белков хлоропласта. Про него известно, что это самый распространенный белок в мире.
Хотя собственно реакция фиксации углерода не требует затраты энергии, для ее протекания нужен непрерывный приток высокоэнергетического вещества – рибулозодифосфата, с которым и связывается CO2.
Эта регенерация требует участия продуктов световой фазы фотосинтеза – АТФ и НАДФН. Как видно из схемы, а здесь приведена лишь часть промежуточных соединений, из трех молекул CO2, вступивших в реакцию, катализируемую рибулозодифосфаткарбоксилазой, образуется шесть молекул 3-фосфоглицерата, из которых в ходе цикла реакций регенерируется три молекулы рибулозодифосфата, использованные в начале цикла и остается одна молекула трехатомного сахара – глицеральдегид-3-фосфата. В цикле фиксации углерода для связывания одной молекулы затрачивается три молекулы АТФ и две молекулы НАДФН.
Глицеральдегид-3-фосфат, образовавшейся при фиксации CO2, является ключевым промежуточным продуктом гликолиза. В строме из него могут образоваться жирные кислоты, аминокислоты, крахмал, в цитоплазме он быстро превращается в результате обратного протекания некоторых реакций гликолиза в производные глюкозы и фруктозы, из которых образуется сахароза.
Если объединить реакции световой и темновой фазы, исключив промежуточные этапы, то получим всем хорошо известное суммарное уравнение процесса фотосинтеза:
6CO2 + 6H2O = C6H12O6 + 6O2.
Как известно, при полном окислении одного моля глюкозы выделяется 686 ккал энергии. Если же посчитать всю энергию, затраченную на синтез этого моля, то получается, что для этого требуется от 1968 до 3456 ккал в зависимости от длины волны поглощаемого света, т.е. эффективность работы фотосинтеза колеблется от 20 до 35%.
Значение фотосинтеза.
Благодаря фотосинтезу, ежегодно из атмосферы поглощаются миллиарды тонн углекислого газа, выделяются миллиарды тонн кислорода; фотосинтез является основным источником образования органических веществ. Из кислорода образуется озоновый слой, защищающий живые организмы от коротковолновой ультрафиолетовой радиации.
При фотосинтезе зеленый лист использует лишь около 1% падающей на него солнечной энергии, продуктивность составляет около 1 г органического вещества на 1 м2 поверхности в час.
Хемосинтез
Синтез органических соединений из углекислого газа и воды, осуществляемый не за счет энергии света, а за счет энергии окисления неорганических веществ, называется хемосинтезом. К хемосинтезирующим организмам относятся некоторые виды бактерий.
Нитрифицирующие бактерии окисляют аммиак до азотистой, а затем до азотной кислоты (NH3 → HNO2 → HNO3).
Железобактерии превращают закисное железо в окисное (Fe2+ → Fe3+).
Серобактерии окисляют сероводород до серы или серной кислоты (H2S + ½O2 → S + H2O, H2S + 2O2 → H2SO4).
В результате реакций окисления неорганических веществ выделяется энергия, которая запасается бактериями в форме макроэргических связей АТФ. АТФ используется для синтеза органических веществ, который проходит аналогично реакциям темновой фазы фотосинтеза.
Хемосинтезирующие бактерии способствуют накоплению в почве минеральных веществ, улучшают плодородие почвы, способствуют очистке сточных вод и др.
Основные понятия генетики живого
Всем хорошо известна способность родительских форм передавать при размножении свои признаки потомству.
Это свойство живых организмов – наследственность, консервативно, оно сохраняет уже возникшие черты и свойства организмов на протяжении многих поколений.
Из предыдущего материала мы уже знаем, что генетическая, или наследственная информация об организме закодирована в ДНК, но она реализуется в определенных условиях внешней среды. Очевидно, что различия в условиях среды накладывают свой отпечаток на особенности развития особи. Все это заставляет рассматривать развитие организма как следствие действия двух главнейших факторов – реализации генетической программы, т.е. действия генотипа, и влияния на особь со стороны факторов среды. Как вы помните,–генотипом называют совокупность всех генов, полученный организмом от его родителей. В генотипе возможны мутации – наследственные изменения, которые приводят к увеличению или уменьшению количества генетического материала, к изменению нуклеотидов или их последовательности. Путем мутаций в генотип включаются новые мутантные гены, которых не было у родителей. Каждый организм имеет свой фенотип, – т.е. комплекс внешних и внутренних признаков организма, таких, как форма, размеры, окраска, химический состав, поведение, биохимические, микроскопические и макроскопические особенности
Любой организм характеризуется совокупностью большого числа признаков и свойств. Признак, или свойство, - единица морфологической, физиологической или биохимической дискретности организма. Условно все признаки можно разделить на качественные и количественные. Первая группа признаков характеризуется тем, что они четко и непосредственно отличаются друг от друга (гладкая или морщинистая форма семян, белая или красная окраска цветков и т.д.), вторая же группа признаков не имеет такого четкого разграничения и может устанавливаться только путем количественного определения (измерения, взвешивания и т.д.).
Деление признаков на качественные и количественные условно, внешние условия, в которых развивается организм, никогда не бывают постоянными, поэтому один и тот же признак выражается в различных величинах (модификациях) но качественные признаки более жестко контролируются генами. Они обладают большей устойчивостью, развитие их относительно меньше зависит от внешних условий и поэтому носит прерывистый характер. Количественные признаки менее устойчивы, развитие их сильно зависит от внешних условий и поэтому носит непрерывный характер.
Формирование признака – это цепь процессов, идущая от гена через мРНК, полипептид и фермент, - протекает нормально только в том случае, если в распоряжении клетки имеются все необходимые исходные вещества, надлежащий источник энергии и подходящие условия для реакций. Таким образом, среда должна обеспечивать условия, необходимые для формирования признака. Например, картофель, помещенный в подвал, не образует зеленых пластид, хотя гены для этого имеются. На свету побеги, образованные таким же картофелем, зеленеют. Синтез хлорофилла зависит, таким образом, не только от соответствующих ферментов, но и от внешнего фактора – света. Гены определяют норму реакции, а от внешней среды зависит, какой вариант в пределах этой нормы реакции реализуется в данном случае.
Мы с вами уже рассмотрели материальные основы хранения, передачи и реализации генетической информации на уровне клетки, сейчас же, после всех приведенных ранее пояснений, приступим к рассмотрению закономерностей наследования на уровне организма, т.е. закономерностей наследования признаков.
И начнем с качественных признаков, которые, как мы уже говорили, мало зависят от внешних условий и жестко контролируются генами.
Основным методом изучения закономерностей наследования признаков на уровне организма является гибридологический метод. В основе этого метода лежит скрещивание (гибридизация) организмов, отличающихся друг от друга по одному или нескольким признакам с последующим, в т.ч. математическим, анализом потомства. В результате скрещивания таких организмов получаются гибридные организмы, или гибриды. Скрещивания, в которых родительские формы отличаются по одной паре признаков, называются моногибридными, при различии по двум парам признаков – дигибридными, а если число признаков больше – полигибридными.
Кроме гибридологического метода, в генетике используют: генеалогический — составление и анализ родословных; цитогенетический — изучение хромосом; близнецовый — изучение близнецов; популяционно-статистический метод — изучение генетической структуры популяций.
При генетическом анализе для записи схем скрещивания пользуются определенными правилами. Родительские формы обозначаются буквой P, женский пол – знаком ♀, мужской - ♂, скрещивание - ×, гибридные поколения – буквой F с соответствующими цифровыми индексами. В генетических работах для обозначения наследственных факторов принято буквенное обозначение. Доминантные гены обозначают заглавными, а соответствующие им рецессивные гены – строчными буквами алфавита. Если доминантный ген желтой окраски семядоли гороха обозначить буквой А, то рецессивный ген зеленой окраски должен быть обозначен буквой а.

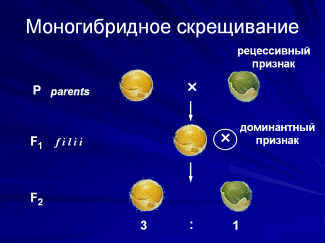
Посмотрим, что происходит при моногибридном скрещивании. Если скрестить растения гороха, имеющего красные цветки с растениями с белыми цветками, то все гибриды первого поколения имеют цветки красного цвета. Признак, проявляющийся у гибридов первого поколения, и ген, ответственный за этот признак, называют доминантным, а не проявляющийся признак и его ген – рецессивным.
Подавление у гибридных организмов одних признаков другими называется доминированием.
Большое число наблюдений и специально поставленные опыты показывают, что доминирование – сложное явление. Оно может изменяться под влиянием внешних условий, возраста, пола, особенностей самого организма, а также других наследственных факторов.
Так у львиного зева гибриды первого поколения от скрещивания красноцветковых растений с белоцветковыми при выращивании на полном свету и при низкой температуре имеют красную окраску цветов, при выращивании в условиях затенения и повышенной температуры цветут белыми цветками, а при промежуточных условиях дают розовые цветки.
У некоторых пород овец проявление признака рогатости зависит от пола: гибридные самцы имеют рога, а у гибридных по этому признаку самок они отсутствуют.
У человека плешивость оказывается доминантным признаком у мужчин и рецессивным – у женщин.
Если вернуться к растениям гороха и получить гибриды второго поколения, произведя скрещивание гибридов первого поколения (в случае гороха это достигается путем самоопыления), то в нем мы уже не будем наблюдать единообразия: часть растений будет иметь один, часть – другой признак исходной родительской пары, причем в распределении доминантных и рецессивных признаков у гибридов второго поколения наблюдается определенная закономерность.
Рассмотрим это на данных, полученных Г.Менделем в его опытах по скрещиванию гибридов первого поколения гороха.

Анализ данных таблицы позволил сделать следующие выводы:
единообразия гибридов во втором поколении не наблюдается: часть гибридов несет один (доминантный), часть — другой (рецессивный) признак из альтернативной пары;
количество гибридов, несущих доминантный признак, приблизительно в три раза больше, чем гибридов, несущих рецессивный признак;
рецессивный признак у гибридов первого поколения не исчезает, а лишь подавляется и проявляется во втором гибридном поколении.
Закономерность в распределении доминантных и рецессивных признаков у гибридов второго поколения в кратном отношении 3:1 называют правилом расщепления, а само явление появления разных типов потомков у внешне одинаковых родителей называется расщеплением.
Гибридные растения первого поколения развиваются в результате слияния гамет с доминантным геном А от красноцветковой родительской формы и с рецессивным геном а от белоцветковой. Поэтому они одновременно имеют и ген красной и ген белой окраски цветков. Так как ген красной окраски доминирует над геном белой, то все гибриды первого поколения оказываются красноцветковыми.
Гибриды первого поколения, однородные с красноцветковыми по фенотипу, в своем генотипе несут гены, обуславливающие развитие разнородных по окраске цветков – красных и белых.
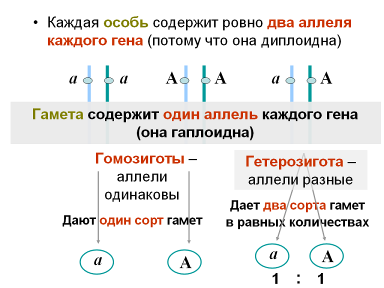
Цитологические основы моногибридного скрещивания вытекают из правил поведения хромосом в мейозе и слияния гамет. При образовании гамет любая из них может получить или доминантный ген А, или рецессивный ген а. Соединение гамет с генами А и а в гибридном организме не приводит к смешиванию или слиянию генов. Гены А и а в гаметах, образуемых гибридными организмами первого поколения, остаются такими же отдельностями, какими они были у исходных родительских форм. В этом заключается чистота гамет в отношении одной пары аллельных генов. Гены одной пары признаков находятся в одинаковых точках гомологичных хромосом. Такие гены получили название аллельных. Аллель – это форма существования гена. Поскольку каждый аллель контролирует один из пары альтернативных признаков, говорят об аллельных признаках как формах проявления гена.
Благодаря наличию двух аллелей возможны и два состояния организма: гетеро- и гомозиготное. Если организм содержит оба одинаковых аллеля данного гена, то он называется гомозиготным по данному гену (или признаку), а если разных, то гетерозиготным.

Понятие аллельности – одно из важнейших. В генетике оно имеет такое же значение, как понятие валентности в химии. Явления наследственности могут быть поняты и объяснены только на основании представления об аллельности дискретных наследственных единиц – генов.



Другим важным понятием генетики является понятие чистоты гамет, цитологической основой которого является локализация аллелей в разных хромосомах каждой гомологичной пары.
Из понятия чистоты гамет вытекает закон чистоты гамет, утверждающий, что признаки не сливаются, не складываются и не делятся, а сохраняются неизменными, распределяясь между различными потомками.
При гибридологическом анализе и в практической селекции применяют взаимные, анализирующие и возвратные скрещивания.
Взаимными, или реципрокными, называют скрещивания между двумя родительскими формами АА и аа, в одной из которых АА является материнской формой, а в другой – отцовской. Формула реципрокных скрещиваний: ♀АА × ♂аа и ♀аа × ♂АА.
Анализирующими называют такие скрещивания, когда какой-либо организм гибридного поколения скрещивают с рецессивной гомозиготной по этому гену исходной родительской формой.
Возвратными, или насыщающими скрещиваниями (беккроссами) называют скрещивание гибридной особи с одной из родительских форм. Такие скрещивания применяют, когда хотят усилить в гибриде проявление признаков какой-либо родительской формы. Широко используется в селекции.
Посмотрим, что получается при дигибридном скрещивании. Горох с желтыми, гладкими семенами (ААВВ) скрещены с зелеными, морщинистыми семенами (аавв). В первом поколении нет ничего особенного. Все растения с желтыми, гладкими семенами. Во втором же поколении расщепление в отношении 9:3:3:1.
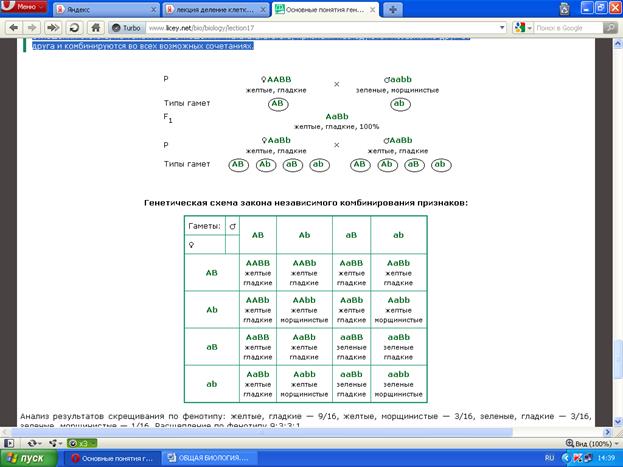
На основании подобных опытов было установлено правило, получившее название закона независимого комбинирования генов, гласящее, что каждая пара аллельных генов (и альтернативных признаков, контролируемых ими) наследуется независимо друг от друга. Цитологические основы закона независимого наследования признаков вытекает из анализа поведения хромосом в мейозе. Попутно отметим, что закон действителен только для генов, локализованных в разных парах гомологичных хромосом.
Если рассмотреть возможные гаметы у организмов первого поколения и всевозможные комбинации гамет с помощью решетки Пеннета, то можно получить16 возможных вариантов зигот, а значит, и потомков. Они распадаются на четыре фенотипических класса в соответствии с вышеприведенным соотношением. Реальное распределения организмов по классам близко к теоретическому, однако редко с ним совпадает, поскольку носит статистический характер, т.к. расщепление определяется вероятностным характером комбинирования в мейозе хромосом, а значит, и заключенных в них генах.
Выводы, полученные при рассмотрении дигибридного скрещивания, позволяют прийти к выводу, что при полигибридном скрещивании для генов, находящихся в разных хромосомах, расщепление во втором поколении будет (3:1)n. Специальные методы статистической обработки позволяют установить достоверность соответствия практически получаемых результатов теоретически ожидаемым.
При формировании представлений о связи между геном и признаком изначально предполагалось, что каждому признаку соответствует особый наследственный фактор, который обуславливает развитие своего признака. Однако такие прямые и однозначные связи гена с признаком на самом деле скорее исключение, чем правило. На самом деле на один признак могут влиять многие гены и, наоборот, один ген часто влияет на многие признаки. Кроме того, действие гена может быть изменено другим геном или условиями внешней среды.


Множественное, или плейотропное, действие генов – это способность гена воздействовать на несколько признаков одновременно. Плейотропия обусловлена тем, что обмен веществ представляет собой сложные метаболические цепи реакций синтеза, превращения и распада. Каждое звено этой цепи контролируется отдельным геном. Мутация любого и них чаще всего затрагивает не один только признак, а несколько и тем самым может влиять на жизнеспособность ее носителей. Причиной этого явления может быть и нарушение синтеза только одного фермента, но участвующего во многих биохимических реакциях.

Примером плейотропного действия гена у человека является болезнь серповидноклеточная анемия. Мутация по этому гену приводит к замене всего лишь одной аминокислоты в молекуле гемоглобина, что изменяет форму эритроцитов (они приобретают серповидную форму вместо двояковогнутого диска) и вызывает нарушения в сердечно-сосудистой, пищеварительной и нервной системах. В гомозиготном состоянии эта мутация является летальной в детском возрасте.
Гены с плейотропным летальным действием приводят к расщеплению, не подчиняются закону расщепления.
Явление, когда за один признак отвечает несколько генов (или аллелей), называется взаимодействием генов.
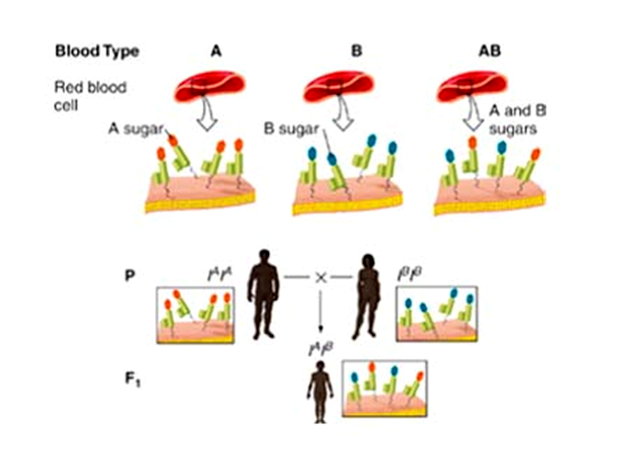
Классическим примером аллельного взаимодействия генов может служить наследование группы АВ крови у человека. Эритроциты людей 1V группы имеют как антигены типа А (детерминируемые геном IА , имеющимся в одной из хромосом клетки), так и антигены типа В (детерминируемые геном IВ, присутствующим в другой гомологичной хромосоме). Таким образом, здесь проявляют свое действие оба аллеля – IА ( в гомозиготном состоянии контролирует II группу крови, группу А) и IВ ( в гомозиготном контролирует III группу В).
Следует отметить, что ген может иметь не два, а большее число аллелей. В данном случае у гена I их три: I0, IА и IВ.



Однако есть гены, имеющие десятки аллелей. Это явление называется множественным аллелизмом, а все аллели одного гена - серией множественных аллелей, из которых каждый диплоидный организм может иметь любые, но только два аллеля. Эти разные аллели одного гена могут проявлять свое действие независимо друг от друга, оказывать модифицирующее влияние или находиться в антагонистических отношениях (доминирование).

Неаллельные взаимодействия генов. Самый распространенный тип взаимодействия генов – это комплементарность, когда гены могут проявить свою активность только при совместном действии, дополняя работу друг друг, а сам по себе ни один из этих генов фенотипического проявления не имеет. Это связано с тем, что синтез большинства сложных соединений представляет собой процесс многоэтапный и каждый этап данного процесса, контролируемый особым ферментом, детерминируется отдельным геном.
Примером такого процесса является наследование окраски цветков у душистого горошка. У этого растения окраска цветка контролируется двумя разными генами, причем каждый доминантный аллель контролирует какой-то один участок цепи биосинтеза пурпурного пигмента, обуславливающего окраску цветка. Поэтому естественно, что синтез пигмента и, следовательно, окраска цветка возможны только при наличии обоих доминантных аллелей. Отсутствие любого из них приводит к отсутствию контролируемого им фермента и блокирует синтез пигмента на одном из этапов.
Естественно, что во взаимодействии могут участвовать несколько генов, возникает полимерия.
Полигены контролируют наследование всех хозяйственно полезных признаков, обуславливающих урожайность и качество урожая сельскохозяйственных растений, продуктивность животных, а также многие важнейшие параметры физической силы, здоровья и умственные способности человека. В значительной степени они подвержены влиянию условий внешней среды, условий выращивания растений и животных, воспитания человека.
При полимерии часто наблюдается так называемое явление трансгрессии, сущность которого состоит в том, что при скрещивании организмов, отличающихся друг от друга по количественному выражению определенного признака, в гибридных потомствах появляются устойчивые (константные) формы с более сильным выражением соответствующего признака, чем это было у обеих родительских форм. Это происходит, когда одна или обе родительские формы не обладают крайней степенью выражения признака, которое может дать данная генетическая система, и, следовательно, в разных локусах хромосом они имеют доминантные и рецессивные аллели. Так, скрещивание ААВВсс × ааввСС в F1 дает тригетерозиготу АаВвСс, а в F2 возникает ряд форм в пределах от ААВВСС до ааввсс. Как видно, расщепление в F2 имеет размах изменчивости выше, чем у обеих родительских форм. Следовательно, при трансгрессиях в гибридном организме объединяются генотипы, дополняющие друг друга.
Из всего сказанного ясно, что проявление большей части признаков не является результатом строго однозначной детерминации признака единственным наследственным фактором, а представляет собой результат влияния целого комплекса взаимодействующих генов и условий внешней среды на формирование каждого конкретного признака.
Мы уже говорили, что закон независимого комбинирования генов действителен только для генов, локализованных в разных парах гомологичных хромосом. Поскольку гомологичных хромосом в организме ограниченное малым числом количество, то ясно, что этому закону могут подчиняться лишь малое число генов. Как же наследуются основная масса генов, число которых в несколько порядков больше числа хромосом?
Благодаря мейозу организм всегда получает от родителей одну из гомологичных хромосом, следовательно, гены, находящиеся в одной хромосоме, передаются потомкам от родителей вместе, образуя группу сцепления. Сцепление может быть полным. Так, Морганом было проведено анализирующее скрещивание дигетерозиготных самцов (серое тело и нормальные крылья) с самками, рецессивными по обоим признакам (черное тело и зачаточные крылья). В результате были получены потомки только родительского типа, т.е. серые с зачаточными крыльями и черные с нормальными крыльями в отношении 1:1. Таким образом, здесь наблюдалось полное сцепление признаков, но не происходило их перемешивание.
Однако это явление полного сцепления скорее исключение, чем правило. Чаще наблюдается появление потомков, несущих как признаки отца, так и признаки матери, но при этом не подчиняющихся закону независимого комбинирования генов. Это так называемое частичное сцепления. Причиной рекомбинации сцепленных генов является кроссинговер, приводящий к обмену участками родительских хромосом и образованию новых рекомбинантных хромосом, содержащих гены как отцовских, так и материнских хромосом.
Гаметы с хромосомами, претерпевшими кроссинговер, называются кроссоверными, а гаметы с хромосомами, образованными без кроссинговера, - некроссоверными. Соответственно этому и особи, возникшие с участием кроссоверных гамет, называются кроссоверными, или рекомбинантными.
Разберем это явление на примере скрещивания двух линий кукурузы, различающихся по окраске эндосперма и консистенции алейроного слоя. Одна линия имеет в гомозиготном состоянии доминантные гены C и S, контролирующие образование окрашенного эндосперма и гладкого алейрона, а другая – их рецессивные аллели c и s, определяющие развитие неокрашенного эндосперма и морщинистого алейрона. Гибридные растения этого скрещивания имеют окрашенный эндосперм и гладкий алейрон. Такие растения должны были бы при независимом комбинировании генов образовать в равном количестве четыре типа гамет: CS, Cs, cS, cs. При анализирующем скрещивании можно было бы ожидать расщепление в отношении 1C-S : 1C-ss : 1cc-S : 1cc-ss. В действительности же 96,4% зерен имеют признаки, свойственные исходным родительским линиям (48,2% окрашенных гладких и 48,2% неокрашенных морщинистых), и только 3,6% зерен имеют новое сочетание признаков. Результаты такого скрещивания можно объяснить только сцеплением генов с последующим кроссинговером.
Для установления принадлежности гена к той или иной группе сцепления проводят скрещивания, учитывая уже имеющиеся данные по генам, положение которых в группах сцепления было определено ранее.
Поскольку гены в хромосомах расположены линейно, то, определяя частоту появления новых сочетаний генов (групп сцепления), можно определить расстояние между генами. За единицу расстояния между генами, называемую морганидой, принят один процент появления новых сочетаний этих генов. Проведя измерения расстояния между генами путем учета процента кроссоверных потомком, можно составить генетические карты хромосом, т.е. определить относительное положение генов в одной группе сцепления.
Мы уже неоднократно говорили, что одной из основных особенностей хромосомного набора любой соматической клетки организма является парность хромосом. Однако это не совсем так. У большинства животных и раздельнополых растений в хромосомном наборе мужской и женской особей хромосомы одной из пар довольно сильно отличаются друг от друга или одна из хромосом представлена в единственном числе. С этими хромосомами связано определение пола и их называют половыми хромосомами. Все остальные хромосомы называют аутосомами. Хромосому, определяющую у ряда организмов мужской пол, называют Y-хромосомой, а непарную ей хромосому – X-хромосомой.
У всех млекопитающих и большинства других видов сочетание XX определяет женский пол, XY – мужской. У птиц и бабочек – обратное определение. У кузнечиков и клопов – XX – женский пол, а XO – мужской, у моли – опять обратное этому определение пола.