Жизнь – это открытая система, развивающаяся на основе матричного автокатализа под влиянием внешнего потока энергии, ограниченная по веществу и использующая циклы реакций.
Приведенное здесь определение жизни – одно из многих, просто потому, что оно больше нравиться мне.
Целостная живая система образуется лишь в результате соединения составных элементов в порядке, который сложился в процессе эволюции.
Целостной живой системе присущи следующие качества:
1. Единство химического состава. Хотя в состав живых систем входят те же химические элементы, что и в объекты неживой природы, соотношение различных элементов в живом и неживом неодинаково. В живых организмах ~ 98% химического состава приходится на шесть элементов: кислород (~62%), углерод (~20 %),водород (~10%), азот (~3%), кальций (~2,5%), фосфор (~1,0 %). Кроме того, живые системы содержат совокупность сложных полимеров (в основном белки, нуклеиновые кислоты, ферменты и т.д.), которые неживым системам не присущи.
2. Открытость живых систем. Живые системы – открытые системы. Живые системы используют внешние источники энергии в виде пищи, света и т.п. Через них проходят потоки веществ и энергии, благодаря чему в системах осуществляется обмен веществ - метаболизм. Основа метаболизма – анаболизм (ассимиляция), то есть синтез веществ, и катаболизм (диссимиляция), то есть распад сложных веществ на простые с выделением энергии, которая используется для биосинтеза.
3. Живые системы – самоуправляющиеся, саморегулирующиеся, самоорганизующиеся системы. Для пояснения этого утверждения дадим определения саморегуляции и самоорганизации.
Саморегуляция – свойство живых систем автоматически устанавливать и поддерживать на определенном уровне те или иные физиологические (или другие) показатели системы. Самоорганизация – свойство живой системы приспособляться к изменяющимся условиям за счет изменения структуры своей системы управления. При саморегуляции и самоорганизации управляющие факторы воздействуют на систему не извне, а возникают в ней самой в процессе переработки информации, которой живая система обменивается с внешней средой. Это означает, что живые системы – самоуправляющиеся системы.
4. Живые системы – самовоспроизводящиеся системы. Живые системы существуют конечное время. Поддержание жизни связано с самовоспроизведением, благодаря чему живое существо воспроизводит себе подобных.
5. Изменчивость живых систем. Изменчивость связана с приобретением организмом новых признаков и свойств. Это явление противоположно наследственности и играет роль в процессе отбора организмов, наиболее приспособленных к конкретным условиям.
6. Способность к росту и развитию. Рост - увеличение в размерах и массе с сохранением общих черт строения; рост сопровождается развитием то есть возникновением новых черт и качеств. Развитие может быть индивидуальным (онтогенез), когда последовательно проявляются все свойства организма, и историческим, которое сопровождается образованием новых видов и прогрессивным усложнением живой системы (филогенез).
7. Раздражимость живых систем. Раздражимость - неотъемлемая черта всего живого. Раздражимость связана с передачей информации из внешней среды к живой системе и проявляется в виде реакций системы на внешние воздействия.
8. Целостность и дискретность. Живая система дискретна, так как состоит из отдельных, но взаимодействующих между собой частей, которые в свою очередь также являются живыми системами. Например: организм состоит из клеток, являющихся живыми системами; биоценоз состоит из совокупностей различных видов, которые также являются живыми системами. С дискретностью связаны различные уровни организации живых систем; о чем будет сказано ниже. Вместе с тем живая система целостна, поскольку входящие в нее элементы обеспечивают выполнение своих функций не самостоятельно, а во взаимосвязи с другими элементами системы.
Специфика живого заключается в том, что ни один из перечисленных признаков (а их число составляет по данным разных ученых до 20-30) не является самым главным, определяющим для того, чтобы систему можно назвать целостной живой системой. Только наличие всех этих признаков вместе взятых позволяет провести границу между живым и неживым в природе. Единственный способ дать определение живому – перечислить основные свойства живых систем.
В начале лекции было упомянуто, что в организации живого можно выделить несколько уровней.
Если это утверждение конкретизировать, то мы получим следующую картину:
1. Молекулярно-генетический
2. Клеточный
3. Тканевый
4. Органный
5. Организменный
6. Популяционно-видовой
7. Биогеоценотический
8. Биосферный
За исключением двух последних пунктов в этом перечне, которые будут рассмотрены в курсе «Общей экологии», в этом порядке будет нами и рассмотрен курс «Биологии».
Химия жизни
Структурная и функциональная организация живого организма немыслима без постоянно протекающих в ней многочисленных химических реакций, в которых участвуют десятки элементов и сотни простых и сложных соединений.
По химическому составу живые организмы могут значительно отличаться друг от друга, но элементы, входящие в их состав, одинаковы. Хотя в составе живых организмов обнаружено около 70 элементов, но лишь 27 из них имеют вполне установленное значение и встречаются в живых организмах постоянно. Из 31 химического элемента, в наибольшем количестве содержащимся в живых организмов только 5 имеют порядковый номер более 34. Более того, соотношение этих элементов в живых организмах совсем иное, чем в земной коре. Интересно отметить, что 8 из 9 элементов, содержащихся в живом организме в наибольших количествах, входят в число десяти элементов, которые в наибольших количествах присутствуют также и в морской воде, что косвенно подтверждает морское происхождение жизни.

В живых организмах в наибольшем количестве встречаются водород, азот, углерод, кислород, на долю которых приходится более 98% общей массы, тогда как в земной коре их доля значительно меньше.

Химические свойства живых организмов в значительной степени зависят от углерода, на долю которого приходится более половины их сухого вещества. Углерод способен образовывать ковалентные связи с большим количеством элементов, но наиболее важное в биологии значение имеет способность углерода к химическим связям друг с другом,

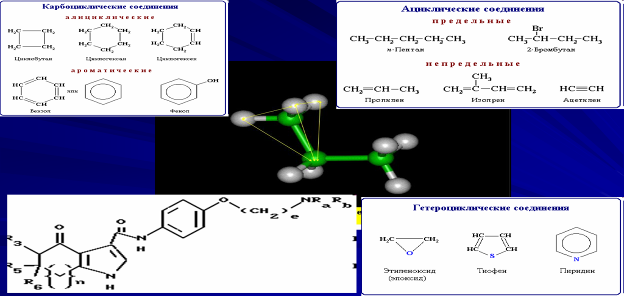 что позволяет формироваться множеству разнообразных структур: линейным и разветвленным цепям, циклическим и сетчатым структурам, а также их комбинации.
что позволяет формироваться множеству разнообразных структур: линейным и разветвленным цепям, циклическим и сетчатым структурам, а также их комбинации.
Все эти структуры лежат в основе скелетов многочисленных органических молекул. Четыре ковалентные одинарные связи углерода располагаются в пространстве в виде тетраэдра, что позволяет образовывать разнообразные трехмерные структуры, на что не способен ни один другой элемент. Трехмерная структура органических молекул играет исключительно важную роль во многих биохимических процессах, а свойство атомов водорода в углеводородной цепи быть замещенным различными функциональными группами, придает большинству биомолекул полифункциональные свойства.
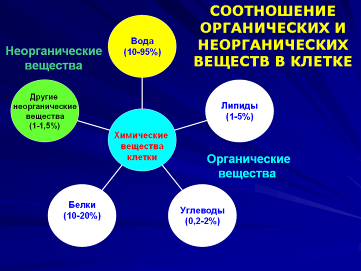
Наибольшую часть (70-97%) массы живых организмов составляет вода. Лишь несколько процентов сухого вещества живого организма приходится на долю неорганических солей и других минеральных веществ, а все остальное приходится на органические соединения, представленные в основном четырьмя видами молекул: белками, нуклеиновыми кислотами, углеводами и жирами.
Если мы рассмотрим химический состав этих молекул, то увидим, что строение этих сложных структур основано на весьма простых принципах. В качестве строительных блоков, из которых они состоят, используются простые молекулы, число которых невелико, и они имеют одно и то же строение у всех видов организмов.
Молекулы всех белков, представляющие собой длинные цепочки, построены всего лишь из 20 разных аминокислот, расположенных в той или иной последовательности. Аналогичным образом длинные, напоминающие цепи, молекулы нуклеиновых кислот у всех организмов построены из небольшого числа нуклеотидов. Белки и нуклеиновые кислоты являются информационными макромолекулами: каждый белок и каждая нуклеиновая кислота несут определенную информацию, закодированную в последовательности строительных блоков. Полисахариды также состоят из большого количества строительных блоков одного-двух типов.
Таким образом, более 90% сухого органического вещества в живых организмах составляют тысячи разнообразных
молекул, построенных всего лишь из трех-четырех десятков различных видов простых органических молекул.
Белок (протеин, полипептид) — высокомолекулярное органическое вещество, состоящее из соединённых в линейную цепочку пептидной связью 20 альфа(L)-аминокислот.
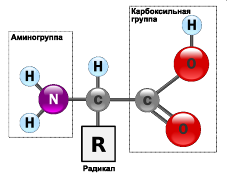
Все они содержат карбоксильную группу и аминогруппу, которые связаны с одним и тем же атомом углерода. Аминокислоты отличаются друг от друга строением только одной части молекулы, а именно боковой группы, которая у разных аминокислот неодинакова по структуре, электрическому заряду и растворимости в воде. Все аминокислоты, кроме одной, имеют асимметричный атом углерода, в результате чего у всех их имеются стереоизомеры. Все аминокислоты, входящие в состав белка, являются L-стереоизомерами.
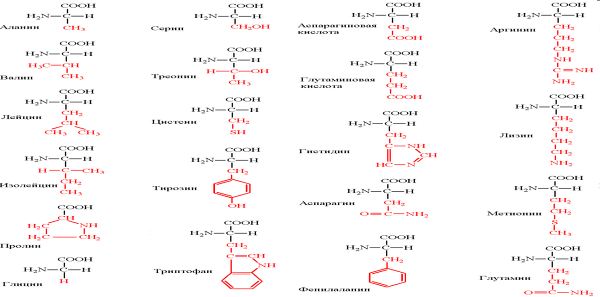
По степени полярности боковой группы все аминокислоты можно расположить в виде непрерывного ряда, начиная с полностью неполярных, или гидрофобных и кончая сильно полярными, или гидрофильными. Являясь бифункциональными соединениями, аминокислоты в водном растворе могут вести себя и как кислоты и как основания, благодаря чему они могут играть роль буферных соединений.
Растения все необходимые аминокислоты синтезируют самостоятельно. Большинство животных, в том числе и человек, потеряли способность синтезировать некоторые аминокислоты, они должны поступать с пищей. Такие аминокислоты называются незаменимыми, их 10.
Наиболее важное для живого свойство аминокислот – способность их молекул соединяться между собой прочными пептидными связями, которые возникают вследствие взаимодействия аминогруппы одной аминокислоты с карбоксильной группой другой с выделением молекулы воды. Так образуется пептид.
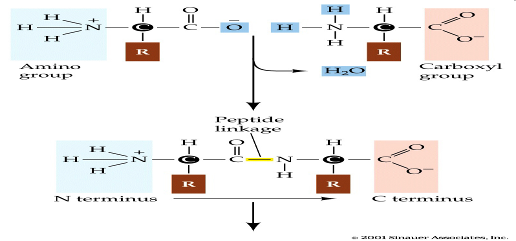
Аминокислотный остаток, находящейся на том конце пептида, где имеется свободная аминогруппа, называется N-концевым остатком, противоположный конец – С-концевой остаток. Дальнейшее присоединение аминокислот с помощью пептидной связи приводит к образованию полипептидной цепочки – первичной структуры белка.
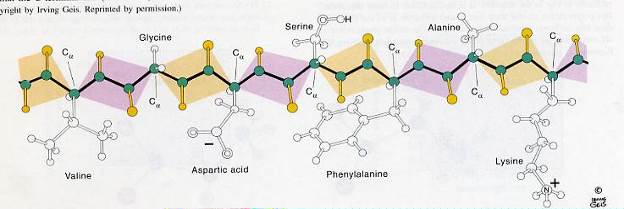
Молекула белка является полипептидом, в состав которого входит от 100 до нескольких тысяч аминокислот. Все молекулы данного индивидуального белка идентичны по аминокислотному составу, последовательности аминокислотных остатков и длине полипептидной цепи.
Характеристика белков не исчерпывается их первичной структурой, поскольку они обладают еще и более высокими уровнями структурной организации. Всякая полипептидная цепь закручивается в спираль из-за образования водородных связей межу CO- и NH-группами каждого пятого остатка аминокислоты, образуя вторичную структуру.
Благодаря наличию функциональных групп в составе полипептидной цепи, эта спираль может свертываться в пространстве определенным образом, вследствие чего возникает характерная для данного типа белка укладка полипептидной цепи – третичная структура.
Белки в своем составе могут иметь несколько полипептидных цепей, образуя четвертичную структуру. Вторичная, третичная и четвертичная структуры при нагревании белка разрушаются, – происходит процесс денатурации белка, разрушения нативной структуры.
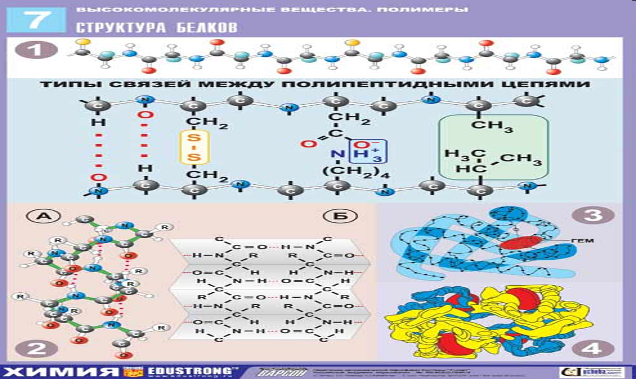
Все белки, в соответствии с их биологическими функциями можно разделить на несколько классов.
Многие белки образуют волокна, навитые друг на друга или уложенные плоским слоем, – они выполняют опорную или защитную функцию, скрепляя биологические структуры и придавая им прочность. Такие белки называются структурными. Главным компонентом хрящей и сухожилий является коллаген, связки содержат эластин, волоса и ногти состоят почти исключительно из кератина.
Есть белки сократительные и двигательные. К ним относятся актин и миозин мышечной ткани, тубулин ресничек и жгутиков.
Белки бываюттранспортные (гемоглобин крови), защитные (иммуноглобулины, фибриноген и тромбин, змеиные яды), регуляторные (некоторые гормоны).
Белки бывают пищевые и запасные. В семенах многих растений запасены пищевые белки, потребляемые на первых стадиях развития зародыша. Пищевые белки – яичный альбумин и казеин. В животных тканях есть ферритин, в котором запасено железо.
Самый многочисленный и наиболее высокоспециализированный класс белков составляют ферменты, или энзимы – белки, обладающие каталитической активностью.
Благодаря ферментам биохимические реакции протекают с огромной скоростью. Скорость ферментативных реакций в десятки тысяч раз (а иногда и в миллионы) выше скорости реакций, идущих с участием неорганических катализаторов. Вещество, на которое оказывает свое действие фермент, называют субстратом.
Активность ферментов определяется их трёхмерной структурой.
Как и все белки, ферменты синтезируются в виде линейной цепочки аминокислот, которая сворачивается определённым образом. Каждая последовательность аминокислот сворачивается особым образом, и получающаяся молекула (белковая глобула) обладает уникальными свойствами.
Чтобы катализировать реакцию, фермент должен связаться с одним или несколькими субстратами. Белковая цепь фермента сворачивается таким образом, что на поверхности глобулы образуется щель, или впадина, где связываются субстраты. Эта область называется сайтом связывания субстрата. Обычно он совпадает с активным центром фермента или находится вблизи него. Такие ферменты, состоящие только из аминокислот, называются простыми ферментами. В то же время некоторые белковые цепи ферментов формируют сайт связывания субстрата лишь после взаимодействия с компонентом небелковой природы (кофактор). Такие ферменты называются сложными. Кофакторы могут быть как неорганическими молекулами (ионы металлов, железо-серные кластеры и др.), так и органическими (например, флавин или гем).
Ферменты, осуществляющие в клетке различные метаболические процессы, например, превращение глюкозы в молочную кислоту в скелетных мышцах, организованы в виде последовательных цепей или систем, в которых они действуют согласованно. В таких реакциях продукт одной является субстратом другой. Такие мультиферментные системы могут включать 15 и более ферментов, действующих в определенной последовательности. В каждой такой системе есть фермент, который определяет скорость реакции всей ферментативной цепочки. Такой фермент способен не только выполнять определенную ферментативную функцию, но и обладает способностью повышать или понижать свою каталитическую активность в ответ на определенные сигналы. У таких ферментов есть сайты связывания малых молекул, они могут быть субстратами или продуктами метаболического пути, в который входит фермент. Наличие или отсутствие малой молекулы в таком сайте приводит к частичному изменению структуры сайта связывания субстрата, что и сказывается на активности фермента. Благодаря действию таких ферментов (аллостерические ферменты), организм способен почти мгновенно приспосабливаться к изменяющимся условиям.
Ферменты обычно проявляют высокую специфичность по отношению к своим субстратам. Это достигается частичной комплементарностью формы, распределения зарядов и гидрофобных областей на молекуле субстрата и в центре связывания субстрата на ферменте. Ферменты демонстрируют высокий уровень стереоспецифичности и селективности.
Для реакций ферментативного катализа характерны: 1) высокая эффективность, 2) строгая избирательность и направленность действия, 3) субстратная специфичность, 4) тонкая и точная регуляция.
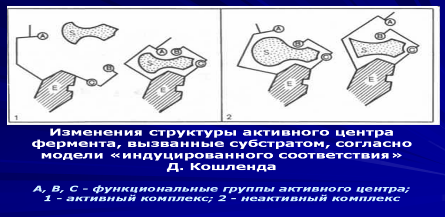
Субстратную и реакционную специфичность реакций ферментативного катализа объясняют гипотезы Э. Фишера (1890 г.) и Д. Кошланда (1959 г.).
Э. Фишер (гипотеза «ключ-замок») предположил, что пространственные конфигурации активного центра фермента и субстрата должны точно соответствовать друг другу. Субстрат сравнивается с «ключом», фермент — с «замком».
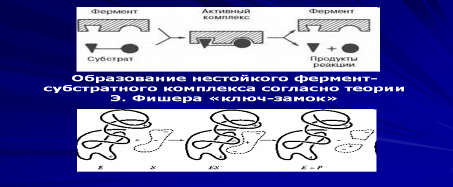
Д. Кошланд (гипотеза «рука-перчатка») предположил, что пространственное соответствие структуры субстрата и активного центра фермента создается лишь в момент их взаимодействия друг с другом. Эту гипотезу еще называют гипотезой индуцированного соответствия.
Скорость ферментативных реакций зависит от: 1) температуры, 2) концентрации фермента, 3) концентрации субстрата, 4) рН. Следует подчеркнуть, что поскольку ферменты являются белками, то их активность наиболее высока при физиологически нормальных условиях.
Большинство ферментов может работать только при температуре от 0 до 40 °С. В этих пределах скорость реакции повышается примерно в 2 раза при повышении температуры на каждые 10 °С. При температуре выше 40 °С белок подвергается денатурации и активность фермента падает. При температуре, близкой к точке замерзания, ферменты инактивируются.
При увеличении количества субстрата скорость ферментативной реакции растет до тех пор, пока количество молекул субстрата не станет равным количеству молекул фермента. При дальнейшем увеличении количества субстрата скорость увеличиваться не будет, так как происходит насыщение активных центров фермента. Увеличение концентрации фермента приводит к усилению каталитической активности, так как в единицу времени преобразованиям подвергается большее количество молекул субстрата.
Для каждого фермента существует оптимальное значение рН, при котором он проявляет максимальную активность (пепсин — 2,0, амилаза слюны — 6,8, липаза поджелудочной железы — 9,0). При более высоких или низких значениях рН активность фермента снижается. При резких сдвигах рН фермент денатурирует.
Скорость работы аллостерических ферментов регулируется веществами, присоединяющимися к аллостерическим центрам. Если эти вещества ускоряют реакцию, они называются активаторами, если тормозят — ингибиторами.
В настоящее время известно несколько десятков тысяч ферментов. Чтобы можно было в них разбираться, принята специальная классификация с систематической номенклатурой. Классификация была предложена Международным союзом биохимии и молекулярной биологии.
По типу катализируемых химических превращений ферменты разделены на 6 классов:
оксиредуктазы (перенос атомов водорода, кислорода или электронов от одного вещества к другому — дегидрогеназа),
трансферазы (перенос метильной, ацильной, фосфатной или аминогруппы от одного вещества к другому — трансаминаза),
гидролазы (реакции гидролиза, при которых из субстрата образуются два продукта — амилаза, липаза),
лиазы (негидролитическое присоединение к субстрату или отщепление от него группы атомов, при этом могут разрываться связи С–С, С–N, С–О, С–S — декарбоксилаза),
изомеразы (внутримолекулярная перестройка — изомераза),
лигазы (соединение двух молекул в результате образования связей С–С, С–N, С–О, С–S — синтетаза).
Классы в свою очередь подразделены на подклассы и подподклассы. В действующей международной классификации каждый фермент имеет определенный шифр, состоящий из четырех чисел, разделенных точками. Первое число — класс, второе — подкласс, третье — подподкласс, четвертое — порядковый номер фермента в данном подподклассе, например, шифр аргиназы — 3.5.3.1.
Другая группа органических соединений живого организма – углеводы, или сахариды. Углеводы являются полигидроксиальдегидами или полигидроксикетонами либо образуют эти вещества в результате гидролиза. Все углеводы делятся на простые (моносахариды) и сложные (полисахариды).
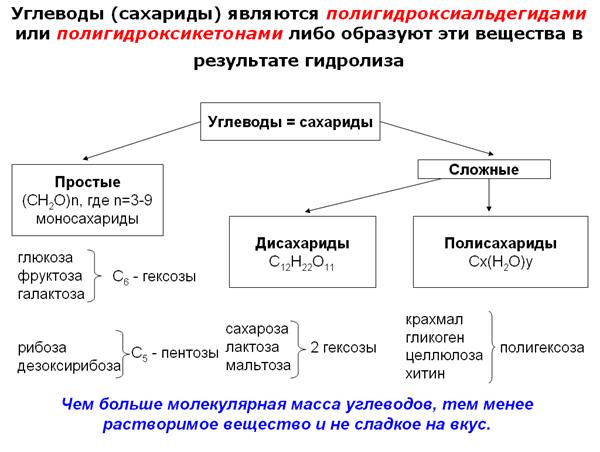
Среди моносахаридов по числу углеродных атомов различают триозы, тетрозы, пентозы, гексозы и гептозы.
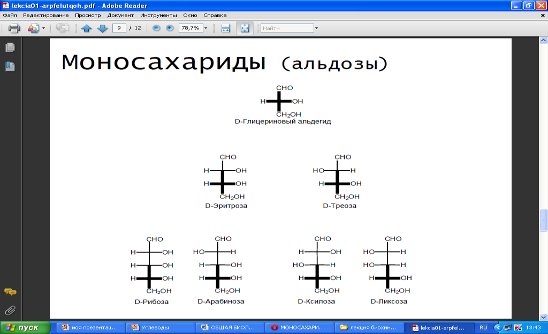
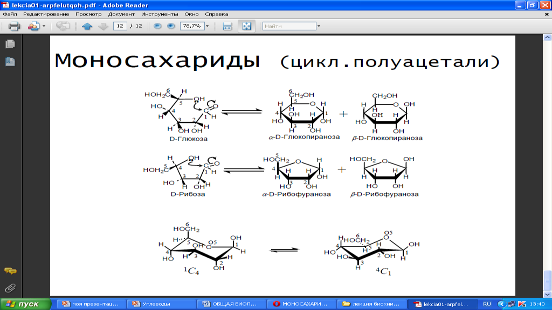
Моносахариды с пятью и более атомами углерода, растворяясь в воде, могут приобретать кольцевую форму. В природе наиболее часто встречаются пентозы и гексозы. Пентозы входят в состав нуклеиновых кислот, гексозы (глюкоза) служат универсальным источником энергии.
Дисахариды состоят из двух моносахаридов, связанных друг с другом ковалентной связью. Мальтоза содержит два остатка глюкозы, связанных друг с другом α(1→4)-гликозидной связью. Лактоза образована из глюкозы и галактозы. Сахароза состоит из остатков глюкозы и фруктозы, соединенных друг с другом через аномерные атомы углерода.

Основная роль углеводов связана с их энергетической функцией. СЛАЙД Полисахариды играют главным образом роль запасных продуктов и легко мобилизируемых источников энергии (крахмал, гликоген), а также используются в качестве строительного материала (целлюлоза, хитин). Полисахариды по ряду причин весьма удобны в качестве запасных питательных веществ: будучи нерастворимы в воде, они не оказывают ни осмотического, ни химического влияния, занимают небольшой объем при большом количестве запасенной энергии и могут быть легко превращены в простые сахара путем гидролиза.
Наиболее важными резервными полисахаридами являются крахмал у растений и гликоген у животных.
Целлюлоза является линейным, неразветвленным гомополисахаридом, состоящим из 10000 и более остатков глюкозы. Полимерные цепи целлюлозы сильно вытянуты и соединяются друг с другом бок о бок, образуя длинные нерастворимые фибриллы.
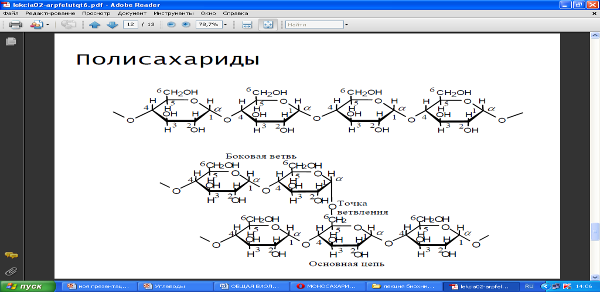
У животных нет ферментов, гидролизующих целлюлозу, поэтому она не может служить пищей для большинства высших животных.
Перед тем как перейти к изучению метаболических процессов, следует рассмотреть еще одну группу биомолекул – липидов. По химическому составу липиды – это сложные эфиры жирных кислот и какого-либо спирта.Жирные кислоты– это небольшие молекулы с длинной цепью, состоящей из атомов углерода (чаще всего 16 или 18) и водорода, и с карбоксильной группой (-СООН). Их углеводородные хвосты гидрофобны, а карбоксильная группа крайне гидрофильна и легко образует эфиры. Иногда в жирных кислотах имеются одна или несколько двойных связей (С = С). В этом случае жирные кислоты, а также содержащие их липиды называются ненасыщенными. Жирные кислоты и липиды, в молекулах которых нет двойных связей, называются насыщенными. Последние образуются путем присоединения дополнительной пары атомов водорода по двойной связи ненасыщенной кислоты.
Ненасыщенные жирные кислоты плавятся при значительно более низких температурах, чем насыщенные. Например, олеиновая кислота (Тпл = 13,4°С) при комнатной температуре бывает жидкой, тогда как пальмитиновая (Тпл = 63,1°С) в этих условиях остается твердой.

Большинство липидов являются сложными эфирами трехатомного спирта глицерина и трех остатков жирных кислот, такие соединения называются триглицеридами. Среди соединений этой группы различают жиры и масла в зависимости от того, остаются ли они твердыми при комнатной температуре (жиры) или находятся в жидком состоянии (масла). В маслах, как правило, присутствует больше ненасыщенных жирных кислот, чем в жирах.
К группе липидов относятся также стероиды, терпены, воска, фосфогликолипиды.
Липиды играют важную роль как источники энергии. При окислении они дают более чем в два раза больше энергии, чем углеводы и белки, и, таким образом, более экономичны как форма хранения запасных веществ. Благодаря низкой теплопроводности липиды выполняют защитные функции, т.е. служат для теплоизоляции организмов. Так как при окислении жиров образуется примерно такое же количество воды, то жиры некоторыми животными (например, верблюдом) используются как источники воды.
Однако главная функция, которую выполняют липиды в организме, это участие в образовании клеточных мембран.
И, наконец, последний по счету, но далеко не самый последний по значимости класс органических веществ живого, который мы рассмотрим достаточно подробно, - это нуклеиновые кислоты.
Нуклеиновые кислоты (НК) – фосфорсодержащие линейные биополимеры живых организмов, обеспечивающие хранение и передачу наследственной информации. Впервые соединения этого плана были найдены в ядрах лейкоцитов, откуда они получили свое название. В природе существует два вида нуклеиновых кислот – дезоксирибонуклеиновые (ДНК) и рибонуклеиновые (РНК). СЛАЙД В состав нуклеиновых кислот входят пуриновое и пиримидиновое основания, пентозный сахар (рибоза или дезоксирибоза) и остаток фосфорной кислоты.
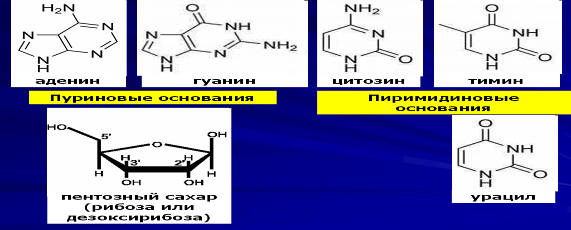
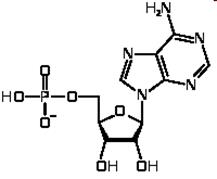
Одно основание, один пентозный сахар и один остаток фосфорной кислоты образуют нуклеотид, который является структурным компонентом (мономером) каждой молекулы НК.
Молекула РНК отличается от ДНК существенно меньшими размерами, она одноцепотчатая, в отличие от двухцепотчатой ДНК, содержит сахар рибозу вместо дезоксирибозы, а вместо основания тимина включает основание урацил.
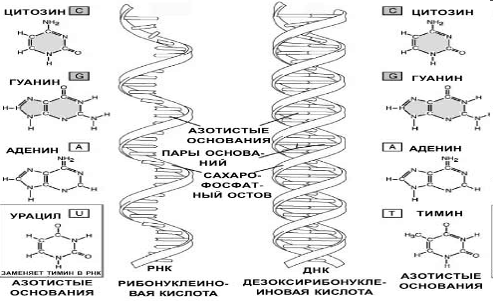
Каждая молекула ДНК или РНК образована всего лишь четырьмя типами нуклеотидов. Последовательно расположенные нуклеотиды в молекулах НК ковалентно связаны друг с другом при помощи фосфатных мостиков. 5’-гидроксильная группа пентозы одного нуклеотида присоединена к 3’-гидроксильной группе пентозы соседнего нуклеотида с помощью фосфодиэфирной связи.
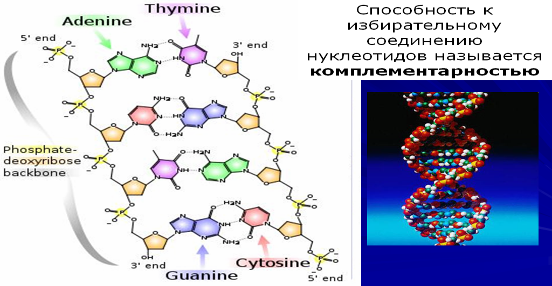
Таким образом, ковалентные остовы НК состоят из монотонно чередующихся фосфатных и пентозных групп, основания же можно рассматривать как боковые группы, присоединенные к остову на равных расстояниях друг от друга. Отметим, что все основания – это почти плоские, гидрофобные молекулы, а сахарофосфатный остов несет заряд, поскольку фосфатные группы являются кислыми и при характерных для клетки рН заряжены отрицательно. Добавим также, что цепи НК обладают определенной полярностью, или направлением, поскольку все межнуклеотидные фосфодиэфирные связи ориентированы вдоль цепи одинаково. Благодаря этой полярности каждая молекула НК имеет 5’-конец и 3’-конец.
Объединяются две полинуклеотидные цепи ДНК в единую молекулу при помощи водородных связей, возникающих между азотистыми основаниями нуклеотидов разных цепей. При этом аденин соединяется только с тимином, а гуанин с цитозином. Благодаря этому свойству последовательность нуклеотидов в одной цепочке определяет их последовательность в другой, т.е. цепи ДНК являются как бы зеркальным отражением одна другой. Такая способность к избирательному соединению нуклеотидов называется комплементарностью, и это свойство лежит в основе образования новых молекул ДНК на базе исходной молекулы, и образования РНК на базе молекулы ДНК.
Принятая в настоящее время модель структуры ДНК выглядит следующим образом. ДНК состоит из двух полинуклеотидных, антипараллельных цепей, закрученных в спираль вокруг общей оси. Антипараллельность цепей проявляется в том, что их 5’-, 3’-межнуклеотидные фосфодиэфирные мостики направлены в противоположные стороны. Гидрофильные остовы цепей, состоящие из остатков дезоксирибозы и фосфорных групп, расположены на внешней стороне двойной спирали и обращены в сторону окружающей ее воды. Гидрофобные основания обеих цепей уложены стопкой внутри двойной спирали, так что практически плоские молекулы оснований сближены между собой и расположены перпендикулярно длинной оси спирали.
Информация записывается вдоль цепи двойной спирали ДНК в форме специфической для данного организма последовательности собранных в тройки азотистых оснований (триплетный код, единый для всего живого).
Исходящая от ДНК информация кодирует аминокислотную последовательность синтезируемых белков. Отрезок ДНК, в котором закодирована информация об одной пептидной цепи белка, представляет собой отдельный ген. Совокупность всех генов конкретного организма составляет его генотип.
У прокариот гены, кодирующие функционально связанные белки, расположены последовательно друг за другом, а вся молекула ДНК – это совокупность генов. У эукариот гены, кодирующие функционально связанные белки, как правило, находятся на разных участках ДНК. В ДНК эукариот помимо уникальных последовательностей нуклеотидов, встречающихся, как правило, один раз (большинство структурных генов, кодирующих мРНК), имеется огромное количество повторяющихся последовательностей – повторов. У высших растений доля уникальных последовательностей не превышает 10-20% всей ДНК, а у млекопитающих на эту последовательность приходится не более 1%.
Большую часть генома занимают интроны – участки ДНК, включенные в состав структурных генов, но не кодирующие их специфический продукт, участок же ДНК, несущий информацию об этом продукте называют экзоном. Самое интересное, многие гены позвоночных, которые были клонированы и детально изучены, содержат большое число интронов (более 50 в гене a-цепи проколлагена).

С точки зрения наличия реально кодирующих индивидуальных последовательностей нуклеотидов в ДНК человек устроен лишь в 6 раз сложнее, чем плодовая муха дрозофила. Для чего нужен высшим организмам такой огромный избыток ДНК не совсем ясно, ясно лишь, что избыток ДНК не является для эукариотических клеток большим неудобством, и что существует довольно слабое давление отбора, направленное на уменьшение общего количества ДНК в клетке за счет сохранения лишь жизненно важных последовательностей.
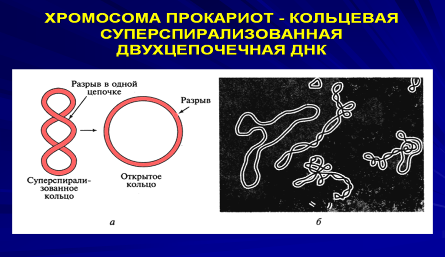
ДНК является главной составной частью хромосомы клетки. Хромосома прокариотической клетки представляет собой одну очень длинную двухцепотчатую молекулу ДНК, собранную в компактное ядерное образование – нуклеоид. У прокариот генетический материал расположен в цитоплазме и не окружен мембраной.
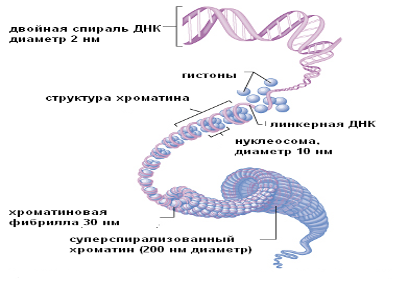
Эукариотические клетки содержат большое число молекул ДНК, каждая из которых, как правило, гораздо длиннее единственной молекулы ДНК прокариот. Молекулы ДНК у эукариот связаны с белками – гистонами, и организованы в хроматин внутри ядра, окруженного сложной двухмембранной системой.
Кроме ядерной ДНК эукариотические клетки содержат небольшое количество ДНК в митохондриях и хлоропластах.

Функция ДНК, как уже отмечалось, состоит в том, что она хранит генетическую информацию, необходимую для кодирования структуры всех белков и всех РНК каждого вида организма, регулирует во времени и пространстве биосинтез компонентов клеток и тканей, определяет деятельность организма в течение его жизненного цикла и обеспечивает индивидуальность данного организма. Как закодирована информация и как она передается, мы рассмотрим в следующих лекциях.
Молекулы РНК по размеру гораздо короче, чем молекулы ДНК, однако их общее количество в большинстве клеток значительно превышает количество ДНК. Как в прокариотических, так и в эукариотических клетках содержится РНК трех основных классов: матричная РНК (мРНК), рибосомная РНК (рРНК) и транспортная РНК (тРНК). Матричная РНК служит матрицей, которая используется рибосомами при переводе генетической информации в аминокислотную последовательность белков. Каждой из 20 аминокислот, входящих в состав белков, соответствует одна или несколько транспортных РНК, которые связывают ее, переносят к рибосомам и служат адаптором при переводе закодированного в мРНК генетического текста в аминокислотную последовательность белка. Роль рибосомной РНК до конца не ясна, известно лишь, что она играет важную роль в структуре и биосинтетической функции рибосом.

Живая клетка
Структурными и функциональными единицами живых организмов являются клетки. Клетка – элементарная живая система, основная структурная и функциональная единица растительного и животного организмов, способная к самообновлению, саморегуляции, самовоспроизведению
Простейшие организмы представляют собой единичные клетки; в отличие от них организм человека содержит не менее 1014 клеток.
Клетки разных биологических видов отличаются и внешним видом и внутренним содержанием, но соблюден некий общий принцип в их устройстве, который позволяет понять, что перед нами именно клетка.
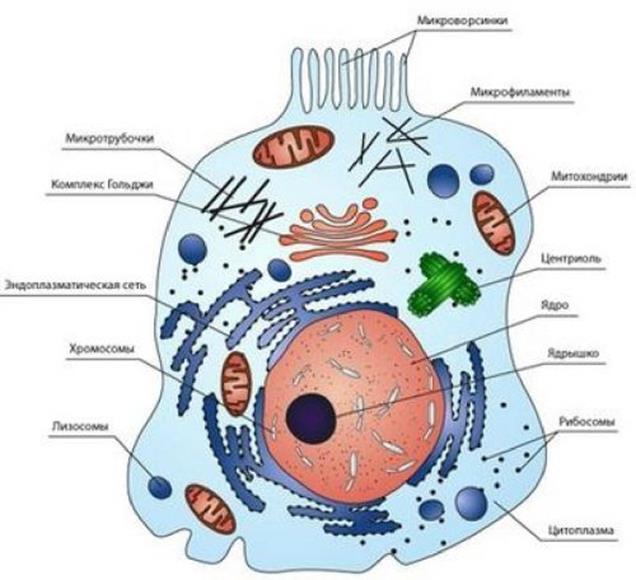
У всякой клетки есть оболочка – мембрана, тонкая пленка, ограничивающая тело клетки в пространстве. Внутри клетка заполнена жидкостью – цитоплазмой. Это самая обычная вода, в которой растворены неорганические и органические вещества. Еще в клетке расположены различные твердые включения, так называемые органоиды или органеллы. Это отдельные крупные молекулы белков и сложные их соединения, молекулярные комплексы.
Все клетки относительно малы. Клетки не могут быть значительно крупнее, чем они есть, просто потому, что в этом случае скорости метаболических процессов могли бы лимитироваться скоростью диффузии молекул веществ внутри клетки, что ограничило бы возможности регуляции метаболизма. Максимальные размеры клеток зависят от основных законов физики, определяющих скорость диффузии молекул, растворенных в воде. Поэтому в наиболее крупных клетках внутренний ее объем разделен на структуры меньших размеров, так называемые компартменты, к которым относятся и клеточные органеллы. Деление на отдельные части всего внутреннего пространства клетки необходимо в значительной мере для того, чтобы облегчить возможность быстрых взаимодействий между специфическими молекулами за счет сокращения пути, который они преодолеют, прежде чем сталкиваются и вступают в реакцию.
Однако, прежде чем мы приступим к подробному рассмотрению строения клетки, надо отметить, что клетки бывают двух типов: прокариотические и эукариотические.
Главное отличие между этими типами клеток состоит в том, что у первых, т.е. прокариотических, отсутствуют структурно оформленное ядро, мембранные органоиды и митоз, у вторых, т.е. эукариотических клеток, все это есть.
Рассмотрим строение прокариотических клеток на примере бактериальной.
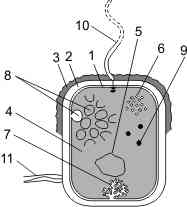
Строение бактериальной клетки:
1 — цитоплазматическая мембрана; 2 — клеточная стенка; 3 — слизистая капсула; 4 — цитоплазма; 5 — хромосомная ДНК; 6 — рибосомы; 7 — мезосома; 8 — фотосинтетические мембраны; 9 — включения; 10 — жгутики; 11 — пили.
Бактериальная клетка ограничена оболочкой. Внутренний слой оболочки представлен цитоплазматической мембраной (1), над которой находится клеточная стенка (2); над клеточной стенкой у многих бактерий — слизистая капсула (3). Строение и функции цитоплазматической мембраны эукариотической и прокариотической клеток не отличаются. Мембрана может образовывать складки, называемые мезосомами (7). Они могут иметь разную форму (мешковидные, трубчатые, пластинчатые и др.).
На поверхности мезосом располагаются ферменты. Клеточная стенка толстая, плотная, жесткая, состоит из муреина (главный компонент) и других органических веществ. Муреин представляет собой правильную сеть из параллельных полисахаридных цепей, сшитых друг с другом короткими белковыми цепочками. В зависимости от особенностей строения клеточной стенки бактерии подразделяются на грамположительные (окрашиваются по Граму) и грамотрицательные (не окрашиваются). У грамотрицательных бактерий стенка тоньше, устроена сложнее и над муреиновым слоем снаружи имеется слой липидов. Внутреннее пространство заполнено цитоплазмой (4).
Генетический материал представлен кольцевыми молекулами ДНК. Эти ДНК можно условно разделить на «хромосомные» и плазмидные. «Хромосомная» ДНК (5) — одна, прикреплена к мембране, содержит несколько тысяч генов, в отличие от хромосомных ДНК эукариот она не линейная, а кольцевая и не связана с белками. Зона, в которой расположена эта ДНК, называется нуклеоидом. Плазмиды — внехромосомные генетические элементы. Представляют собой небольшие кольцевые ДНК, не связаны с белками, не прикреплены к мембране, содержат небольшое число генов. Количество плазмид может быть различным. Наиболее изучены плазмиды, несущие информацию об устойчивости к лекарственным препаратам (R-фактор), принимающие участие в половом процессе (F-фактор). Плазмида, способная объединяться с хромосомой, называется эписомой.
В цитоплазме бактерий находятся рибосомы 70S-типа (6) и включения (9). Как правило, рибосомы собраны в полисомы. Каждая рибосома состоит из малой (30S) и большой субъединиц (50S). Функция рибосом: сборка полипептидной цепочки. Включения могут быть представлены глыбками крахмала, гликогена, волютина, липидными каплями.
У многих бактерий имеются жгутики (10) и пили (фимбрии) (11). Жгутики не ограничены мембраной, имеют волнистую форму и состоят из сферических субъединиц белка флагеллина. Эти субъединицы расположены по спирали и образуют полый цилиндр диаметром 10–20 нм. Жгутик прокариот по своей структуре напоминает одну из микротрубочек эукариотического жгутика. Количество и расположение жгутиков может быть различным. Пили — прямые нитевидные структуры на поверхности бактерий. Они тоньше и короче жгутиков. Представляют собой короткие полые цилиндры из белка пилина. Пили служат для прикрепления бактерий к субстрату и друг к другу. Во время конъюгации образуются особые F-пили, по которым осуществляется передача генетического материала от одной бактериальной клетки к другой.
Эукариотическая клетка характеризуется наличием ядра и изобилием и сложностью строения мембран. Мембраны окружают не только цитоплазму, но и ядро, митохондрии и пластиды, они образуют лабиринт эндоплазматического ретикулума и стопки уплощенных пузырьков аппарата Гольджи. Мембраны окружают лизосомы, пероксисомы и крупные вакуоли. Все эти окруженные мембранами структуры соответствуют определенным компартментам цитоплазмы, предназначенным для тех или иных специализированных метаболических процессов и циклов.
Все клеточные мембраны построены по единому плану. Основой мембраны является двойной слой фосфолипидов с некоторым количеством других липидов, причем липиды повернуты друг к другу своими гидрофобными концами. Липиды, входящие в состав мембранного бислоя, не закреплены жестко, а находятся в непрерывном движении. Однако движение наблюдается в основном в пределах одного слоя, переход молекулы липида из одного слоя в другой – явление редкое.
В жидких слоях мембран находятся специализированные протеиновые комплексы. Липопротеины погружены в липидную фазу и удерживаются гидрофобными связями. Гидрофильные периферические белки удерживаются на внутренней и внешней поверхности мембран электростатическими связями, взаимодействуя с гидрофильными головками липидов.

Мембраны непроницаемы для заряженных молекул, крупных полярных молекул, трудно проницаемы для средних полярных молекул и легко проницаемы для малых молекул, в т.ч. воды.
Каждая клетка окружена мембраной, ограничивающей ее содержимое и позволяющей ей быть до некоторой степени самостоятельной. Клеточная мембрана (плазматическая мембрана) характеризуется избирательной проницаемостью. Это свойство позволяет необходимым питательным веществам и солям проникать внутрь клетки, а лишним продуктам выходить из нее. В то же время клеточная мембрана препятствует проникновению в клетку ненужных веществ из окружающей среды.
С внешней стороны плазматической мембраны многие клетки животных тканей имеют тонкую гибкую клеточную оболочку. На наружной стороне плазматической мембраны содержится большое количество разнообразных полисахаридных, липидных и белковых молекул. На поверхности клетки находится много различных молекул, принимающих и распознающих внешние сигналы. К их числу относятся участки распознания клеток, при помощи которых однотипные клетки узнают друг друга и прикрепляются одна к другой, образуя специфические ткани. На поверхности многих животных клеток имеются рецепторные участки, связывающие различные гормоны. Есть участки распознания чужеродных для данного организма белков и многие другие участки. Таким образом, поверхность многих животных клеток представляет собой поистине сложную мозаику, составленную из различных чувствительных молекулярных антенн, при помощи которых клетки общаются с внешним миром и отвечают на воздействие специфических агентов, присутствующих в окружающей среде.
Внутреннее пространство любой клетки заполнено цитоплазмой, в которой протекает большинство катализируемых ферментами реакций клеточного метаболизма. Все цитоплазматические органеллы погружены в сплошную водную фазу – цитозоль. Цитозоль - это не просто разбавленный водный раствор; его состав весьма сложен, а консистенция приближается к гелю. В цитозоле растворены многие ферменты и ферментные системы, а также другие белки, обеспечивающие связывание, хранение и транспорт питательных веществ, микроэлементов и кислорода. В цитозоле содержится огромное количество небольших биомолекул, коферментов, АТФ и т.п. Все составные части цитозоля поддерживаются в постоянных концентрациях и сбалансированных пропорциях благодаря функционированию систем, обеспечивающих их транспорт через плазматическую мембрану.
Эукариотическая клетка содержит высокоорганизованное, очень сложное ядро, окруженное ядерной оболочкой, состоящей из двух мембран. Через определенные интервалы обе мембраны ядерной оболочки сливаются, образуя отверстия – ядерные поры, через которые происходит обмен различными веществами между ядром и цитоплазмой. Внутри ядра находится ядрышко – место синтеза РНК и начала сборки рибосом. В остальной части ядра находится хроматин (напомним, что хроматин это сложный комплекс ДНК и белка гистона). Хроматин в промежутках между делениями клетки без видимого порядка распределяется по всему ядру, а непосредственно перед делением собирается в хромосомы.
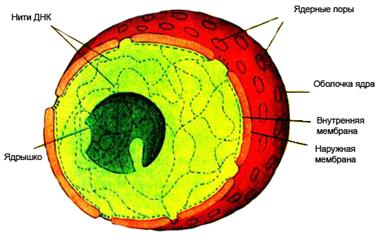
Все клетки живых организмов одного вида содержат строго определенное число хромосом. Каждая из хромосом имеет свою индивидуальную форму и индивидуальное генетическое содержание. Форма и число хромосом организма называется его кариотипом. Сохраняя всю полноту свойственной им генетической информации, хромосомы меняют свою длину в процессе прохождения клеточного цикла, о чем мы будем говорить далее. Характерной чертой строения ядер клеток является парность хромосом. Каждая индивидуальная хромосома имеет в ядре своего гомолога. У человека 46 хромосом составляют 23 пары, у мягкой пшеницы 42 хромосомы составляют 21 пару, у кукурузы имеется 10 пар хромосом. Сущность парности хромосом состоит в том, что один гомолог в паре организм получает от матери, а другой - от отца. ДНК, содержащаяся в одном наборе хромосом, называется геномом.
Весьма заметное место в цитоплазме клеток занимают митохондрии, размеры, внешний вид и число которых может значительно варьировать от клетки к клетки. Каждая митохондрия имеет две мембранные системы. Гладкая внешняя мембрана полностью окружает всю митохондрию. Внутренняя же мембрана образует выступающие внутрь митохондрии складки – кристы. Внутренняя часть митохондрии заполнена гелеобразным веществом – матриксом. В матриксе содержатся рибосомы и митохондриальная ДНК, ответственные за синтез части белков митохондрий. ДНК в митохондриях имеет кольцевое строение.
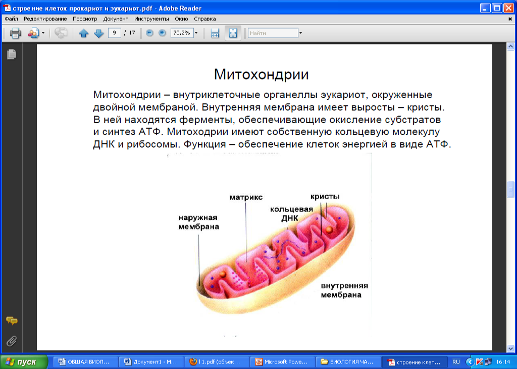
Собственная генетическая система и способность митохондриальной ДНК к репликации, т.е. к самоудвоению, позволяет митохондриям делиться независимо от деления ядра. В клетке они образуются от предсуществующих митохондрий. Продолжительность жизни митохондрий определяется скоординированной деятельностью ядра, цитоплазмы и самих митохондрий.
В митохондриях функционируют системы аэробного дыхания и окислительного фосфорилирования, обеспечивающие энергетические потребности клеток. Во внутренней мембране митохондрий локализованы компоненты электронтранспортной цепи и АТФ-синтетазные комплексы, осуществляющие транспорт электронов и протонов и сопряженный с ними синтез АТФ. В матриксе располагаются ферментные системы цикла Кребса, а также ряд систем синтеза липидов и аминокислот.
В цитоплазме всех клеток имеется очень сложный трехмерный лабиринт мембранных каналов – эндоплазматический ретикулум, многочисленные складки и разветвления которого заполняют всю цитоплазму. Пространство внутри ЭПР, называемое цистернами, используется в качестве каналов, по которым осуществляется транспорт различных веществ. ЭПР делится на два вида – гладкий и шероховатый. Наружная поверхность мембран последнего усеяна рибосомами, тогда как на гладком ЭПР рибосом нет. Белок, синтезированный рибосомами, проталкивается через мембрану внутрь цистерн, откуда в конечном итоге транспортируется к месту назначения.
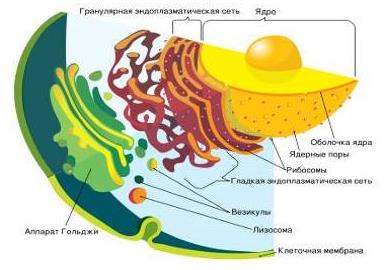
Все клетки содержат характерные скопления окруженных мембранами пузырьков, называемых аппаратом Гольджи. Аппарат Гольджи является продолжением ЭПР в котором в конечном итоге собираются продукты, транспортируемые по цистернам ЭПР, упаковываются в секреторные пузырьки, которые затем перемещаются к внешней плазматической мембране и сливаются с ней. Последующий разрыв пузырька приводит к излиянию его содержимого во внешнюю среду, а сама мембрана пузырька вливается в состав плазматической мембраны, увеличивая ее в размерах.
Лизосомы, – окруженные мембраной сферические пузырьки. Лизосомы содержат много различных ферментов, способных переваривать, т.е. расщеплять путем гидролиза, уже ненужные клеточные белки, полисахариды и липиды. Поскольку такие ферменты могут разрушать и остальное содержимое клетки, они заключены в лизосомы. Белки и другие компоненты, которые необходимо разрушить, избирательно переносятся внутрь лизосом, где подвергаются гидролитическому расщеплению до простейших составных частей, поступающих обратно в цитоплазму.
Еще один тип окруженных мембраной цитоплазматических органелл представляют пероксисомы. Внутри этих органелл заключены ферменты, образующие и использующие перекись водорода, а также каталаза – фермент, разрушающий перекись водорода.
В цитоплазме клеток содержатся многочисленные нити, или филаменты, состоящие из последовательно соединенных друг с другом белковых молекул. Наряду с филаментами ряд клеток, в частности клетки нервной системы животных содержат микротрубочки, состоящие из плотно упакованных белковых молекул, расположенных вокруг полой сердцевины. В нервных клетках пучки таких трубочек участвуют в транспорте веществ из тела клетки к концам клеточных отростков – аксонов. Кроме того, трубочки принимают участие в работе митотического веретена во время деления клеток, играют роль двигательных элементов в ворсинках и жгутиках эукариот.
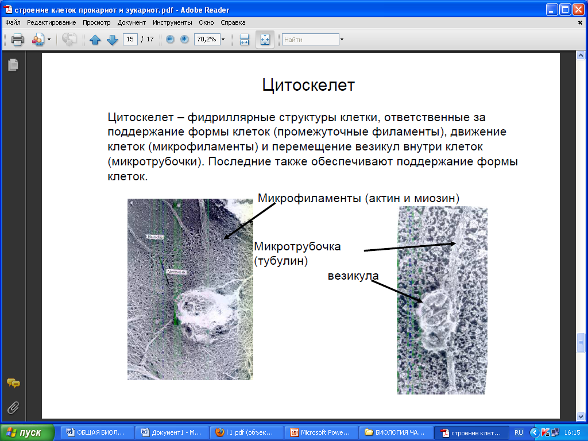
Во многих клетках микрофиламенты различных типов и микротрубочки в совокупности образуют гибкий скелет, называемый цитоскелетом. Цитоскелет придает клеткам их характерный внешний вид и форму, служит местом прикрепления внутриклеточных органелл, фиксируя их местоположение в клетке, а также обеспечивает возможность взаимосвязи между составными частями клетки.
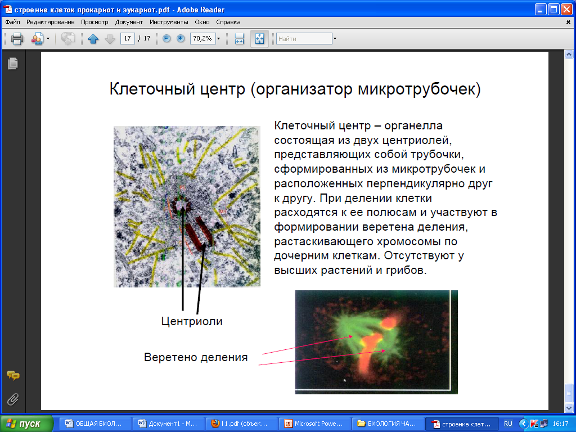
Центриоли - разновидность микротрубочек, представляет собой два очень маленьких тельца цилиндрической формы, расположенных под прямым углом друг к другу. Центриоли относятся к самовоспроизводящимся органоидам цитоплазмы. Их воспроизведение осуществляется путём самосборки из белковых субъединиц.
Во время деления клетки они образуют веретено, вдоль которого выстраиваются хромосомы.
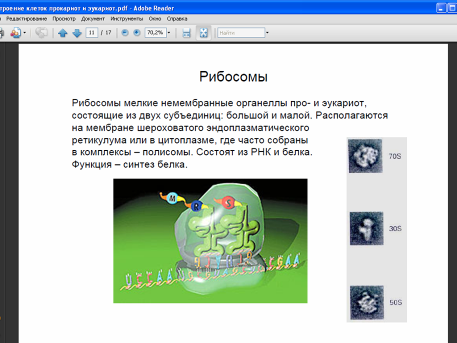
В цитоплазме также содержатся гранулярные элементы, не ограниченные мембранами. Главные из них – это рибосомы, одни из которых находятся в цитоплазме в свободном виде, другие связаны с ЭПР. Рибосомы эукариот крупнее рибосом прокариот, однако, и те и другие выполняют одну и ту же функцию: биосинтез белков из аминокислот.
Другой тип гранулярных элементов в цитоплазме эукариотических клеток – это гранулы гликогена, которые наиболее часто встречаются в клетках печени. Гранулы гликогена выполняют функцию запасных источников энергии, особенно в клетках мышц и печени. В некоторых клетках содержатся жировые капельки, функция которых та же.
Эукариотические клетки высших растений несколько отличаются от клеток высших животных, несмотря на сходство их основных особенностей. Растительные клетки содержат пластиды.
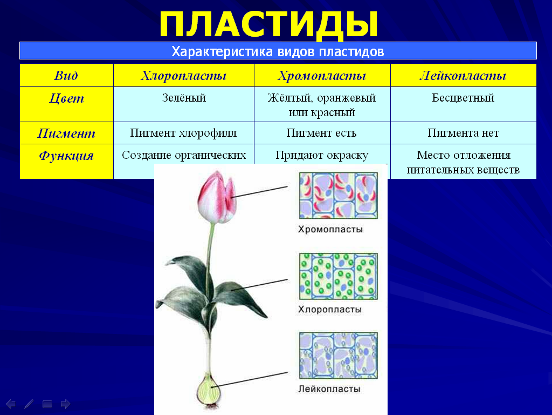
Пластиды – это специализированные органеллы, окруженные двойной мембраной. К самым типичным пластидам, характерным для всех клеток зеленых растений, относятся хлоропласты.
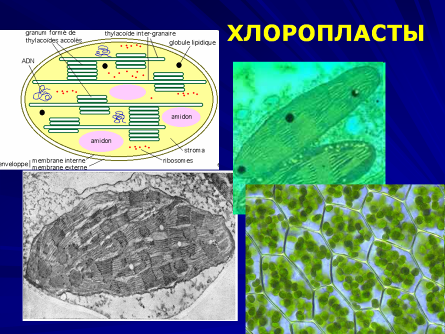
Хлоропласты значительно крупнее митохондрий и могут иметь самую разную форму. Как и другие пластиды, хлоропласты имеют наружную и внутреннюю мембраны. Внутренняя мембрана, ограничивающая внутреннюю гомогенную среду (строму) хлоропласта, в отличие от митохондрий не образует крист и не содержит цепи переноса электронов. Хлоропласты имеют третью тилакоидную мембрану, формирующую группу уплощенных дисковидных мешочков – тилакоидов. Внутренние полости тилакоидов сообщаются между собой, образуя третье внутреннее пространство хлоропласта, называемое тилакоидным пространством. Несколько тилакоидов, лежащих друг над другом, формируют стопку – грану. В мембранах тилакоидов локализованы зеленые (хлорофиллы), желтые и красные (каротиноиды) пигменты, а также компоненты цепи переноса электронов, участвующие в поглощении и использовании энергии света. Биохимические системы синтеза и превращения углеводов функционируют в строме хлоропластов. В ней же откладывается крахмал. Подобно митохондриям хлоропласты тоже содержат ДНК, РНК и рибосомы.
При росте клетки количество хлоропластов увеличивается путем деления. Деление хлоропластов происходит через 6-20 часов и не обязательно совпадает с делением ядра. Хлоропласты являются теми солнечными силовыми станциями, где происходит улавливание солнечной энергии и преобразование ее в энергию химических связей органических веществ.
Среди других пластид следует отметить бесцветные лейкопласты, в которых запасаются крахмал и масла.
Еще одной органеллой, характерной только для растений, является вакуоль.
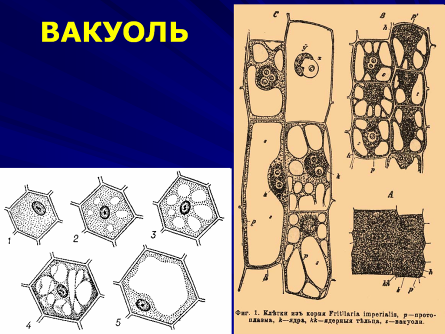
В меристематических клетках вакуоли представлены мелкими пузырьками; для зрелых клеток характерна большая центральная вакуоль. Вакуолярная система растений формируется несколькими путями. Из расширенных цистерн ЭПР образуются провакуоли, слияние которых приводит к возникновению более крупных вакуолей и созданию вакуолярной мембраны – тонопласта, который таким образом является производным ЭПР. Тонопласт может образовывать инвагинации, что приводит к включению в вакуоль участков цитоплазмы. Гидролитические ферменты, содержащиеся в возникшей вакуоли, расщепляют полимеры до низкомолекулярных веществ.
Внутреннее содержание вакуоли, или клеточный сок, имеет сложный состав и включает органические вещества и минеральные соли, углеводы, аминокислоты и белки, которые могут быть вторично использованы в обмене веществ.
Растительный организм не имеет системы выделения, поэтому все продукты метаболизма, а именно, фенолы, танины, алкалоиды, антоцианы и другие продукты обмена выводятся из цитоплазмы клетки в вакуоль и таким путем изолируются. Часто продукты метаболизма агрегируются с образованием кристаллических отложений, это снижает их концентрацию, что позволяет растительной клетке регулировать осмотическое давление.
Вещества доставляются в вакуоль с помощью разнообразных систем транспорта, функционирующих в тонопласте. Важно отметить, что вакуоль может служить местом отложения запасных веществ (алейроновые зерна). Процесс вакуолизации - необходимое условие роста клеток растяжением.
Большинство клеток высших растений полностью окружено клеточной стенкой.
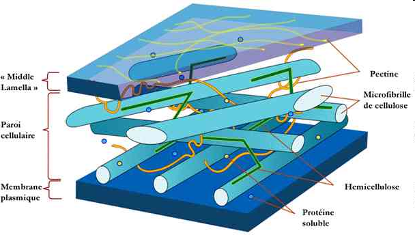
Клеточная стенка сравнительно толста, имеет пористое строение и очень прочна. Она состоит из целлюлозных волокон, склеенных друг с другом сложными полимерными цементирующими веществами. Вода и небольшие молекулы легко проходят через клеточную стенку, которая, однако, предохраняет клетку от набухания и растяжения. Будучи продуктом метаболической активности протопласта, клеточная стенка выполняет функцию защиты содержимого клетки от повреждений и избыточной потери воды, поддерживает форму (за счет тургора) и определяет размер клетки, служит важным компонентом ионного обмена клетки и местом транспорта веществ из клетки. Биогенез клетки играет важную роль в росте и дифференцировке клетки.
Деление клетки
Главные явления в процессах воспроизведения живого – это ауторепродукция наследственных структур и последующее деление клетки. Эти явления направлены на то, чтобы дочерние клетки получили всю полноту генетической информации. В основе ауторепродукции лежат процессы самоудвоения молекул ДНК, без чего нельзя было бы во всей полноте воспроизвести генетическую информацию, свойственную исходной клетке. О механизме самоудвоения ДНК мы поговорим позднее, сейчас же на примере эукариотических клеток рассмотрим вопрос о том, что происходит с генетическим материалом клеток при их делении.
Деление клетки начинается с деления ядра, расхождения дочерних ядер к разным полюсам клетки, затем происходит разделение цитоплазмы.
Способы деления эукариотических клеток: митоз, мейоз, амитоз
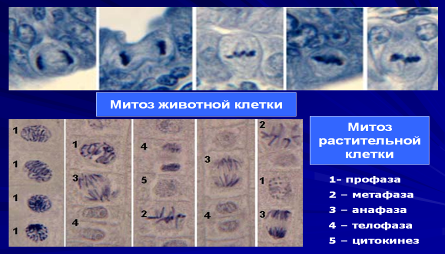
Митоз — основной способ деления эукариотических клеток, при котором сначала происходит удвоение, а затем равномерное распределение между дочерними клетками наследственного материала. Длительность митоза различна у разных клеток, но обычно не превышает 10 минут. Митоз характерен для соматических клеток и позволяет сохранять диплоидное число хромосом в этих клетках во всех последующих поколениях.
Митоз представляет собой непрерывный процесс, в котором выделяют четыре фазы: профазу, метафазу, анафазу и телофазу. Перед митозом происходит подготовка клетки к делению, или интерфаза. Период подготовки клетки к митозу и собственно митоз вместе составляют клеточный (митотический) цикл. Ниже приводится краткая характеристика фаз цикла.

Интерфаза состоит из трех периодов: пресинтетического, или постмитотического, — G1, синтетического — S, постсинтетического, или премитотического, — G2.
Пресинтетический период (2n 2c, где n — число хромосом, с — число молекул ДНК) — Клетка только что поделилась
Идут процессы синтеза белков, жиров, нуклеиновых кислот и углеводов
Синтезируются и развиваются клеточные органеллы (размножаются митохондрии, развивается система ЭПР, синтезируются органоиды движения и пр.)
Клетка выполняет свою физиологическую функцию в организме (например, что-нибудь секретирует)
Запасаются необходимые мономеры (нуклеотиды) для репликации ДНК
Синтезируются ферменты, которые будут осуществлять удвоение генетического материала
Синтетический период (2n 4c) — репликация ДНК.
Постсинтетический (G2) период (2n 4c) — подготовка клетки к митозу, синтез и накопление белков и энергии для предстоящего деления, увеличение количества органоидов, удвоение центриолей.
Клетка перед митозом.
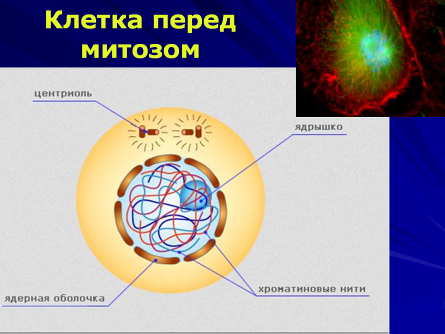
Каких-либо особенностей в клетке не видно, за исключением разделившейся центриоли.
Собственно митоз начинается с конденсации хроматина в хромосомы, которые становятся видными в ядре. Еще раз обращаем внимание на то, что каждая хромосома входит в митоз уже разделенной на две хроматиды. Поэтому в митозе собственно происходит лишь распределение хроматид в дочерние клетки.
Профаза (2n 4c) — демонтаж ядерных мембран, расхождение центриолей к разным полюсам клетки, формирование нитей веретена деления, «исчезновение» ядрышек, конденсация двухроматидных хромосом.
Метафаза (2n 4c) — выстраивание максимально конденсированных двухроматидных хромосом в экваториальной плоскости клетки (метафазная пластинка), прикрепление нитей веретена деления одним концом к центриолям, другим — к центромерам хромосом.
Анафаза (4n 4c) — деление двухроматидных хромосом на хроматиды и расхождение этих сестринских хроматид к противоположным полюсам клетки (при этом хроматиды становятся самостоятельными однохроматидными хромосомами).
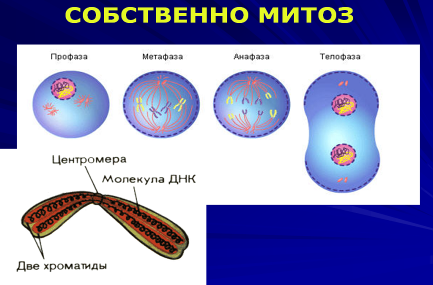
Телофаза (2n 2c в каждой дочерней клетке) — деконденсация хромосом, образование вокруг каждой группы хромосом ядерных мембран, распад нитей веретена деления, появление ядрышка, деление цитоплазмы (цитотомия). Цитотомия в животных клетках происходит за счет борозды деления, в растительных клетках — за счет клеточной пластинки.
Биологическое значение митоза. Образовавшиеся в результате этого способа деления дочерние клетки являются генетически идентичными материнской. Митоз обеспечивает постоянство хромосомного набора в ряду поколений клеток. Лежит в основе таких процессов, как рост, регенерация, бесполое размножение и др.
Существует регулирующий механизм, который позволяет сохранять постоянство объема генетической информации по поколениям организмов. Таким механизмом служит мейоз, который предшествует образованию как женских, так и мужских половых клеток. Мейоз — это особый способ деления эукариотических клеток, в результате которого происходит переход клеток из диплоидного состояния в гаплоидное.
Итак, давайте еще раз вспомним, В чем значение процесса обычного клеточного деления - митоза? Клеток становится больше. При этом все образующиеся клетки генетически однородны!
Многоклеточные организмы развиваются из зиготы именно благодаря митозу
Регенерация и заживление ран происходит за счет митоза
Что значит «генетически однородны»? Допустим, в материнской клетке находится 4 хромосомы
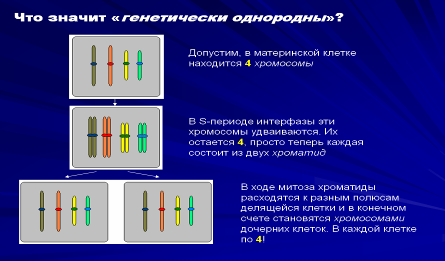
В S-периоде интерфазы эти хромосомы удваиваются. Их остается 4, просто теперь каждая состоит из двух хроматид
В ходе митоза хроматиды расходятся к разным полюсам делящейся клетки и в конечном счете становятся хромосомами дочерних клеток. В каждой клетке по 4!
В ходе митоза хроматиды расходятся к разным полюсам делящейся клетки и в конечном счете становятся хромосомами дочерних клеток. В каждой клетке по 4!
В мейозе все несколько по-другому.. В результате мейоза образуются генетически разнородные клетки
Количество генетического материала в каждой дочерней клетке уменьшается вдвое!
Деление не одно – а два подряд!!
А всего клеток образуется 4 (четыре)!!!
Начинается все практически так же.. Есть клетка. В ней те же 4 хромосомы.. В S-периоде они так же удваиваются и тоже теперь состоят из двух хроматид.. НО!
Мейоз состоит из двух клеточных делений при одном синтезе ДНК.
При первом делении гомологи каждой пары хромосом, имеющие после синтеза ДНК в интерфазе по две хроматиды конъюгируют, т.е. плотно прижимаются друг к друга. Эта конъюгация гомологичных хромосом отличается удивительной точностью, осуществляясь на молекулярном уровне. В процессе этой конъюгации между хроматидами гомологичных хромосом происходит кроссинговер, т.е. обмен гомологичными участками между несестринскими хроматидами.

В дальнейшем между хроматидами возникает отталкивание, и пары сестринских хроматид отделяются друг от друга, оставаясь связанными в центромере и образуя тетрады.
В ооцитах ряда позвоночных, которым свойственен длительный период развития и накопления запасных веществ, процессы мейоза задерживаются на этой стадии на длительное время. У девочек ооциты достигают этой стадии уже на пятом месяце эмбрионального развития и задерживаются на ней вплоть до овуляции. Таким образом, задержка мейоза на этой стадии у женщин в целом составляет период времени от ~ 12 лет (наступление первой овуляции) до ~ 50 лет (окончания периода овуляции).
На следующей стадии мейоза тетрады располагаются по экватору клетки, ядерная оболочка растворяется, к центромерам хромосом прикрепляется нить веретена, и хр
Дата добавления: 2015-11-16; просмотров: 4281;