Глава 6 РОЛЬ ИММУННОЙ СИСТЕМЫ В ПАТОЛОГИИ
6.1. ФУНКЦИОНАЛЬНАЯ ОРГАНИЗАЦИЯ ИММУННОЙ СИСТЕМЫ
6.1.1. Основные понятия
Иммунитет- это общебиологический феномен, суть которого заключается в долговременном самоподдержании внутри отдельного организма баланса между генетически «своим» и«несвоим» в условиях чужеродного окружения. Иммунная системаспециализируется в реализации механизмов данного феномена. Чтобы понять основы науки иммунологии,необходимо изучить защитные механизмы, направленные на устранение «несвоего», и регуляторные процессы, которые формируют внутренний го-меостаз «своего» в организме. Иммунитет может быть врождённым (или неспецифическим) и адаптивным (приобретённым, специфическим). Строго говоря, об иммунитете целесообразно говорить только во втором случае; врождённый иммунитет - это скорее совокупность факторов и механизмов неспецифической резистентности. Отказ одного или нескольких компонентов иммунной системы может приводить к развитию иммунодефицитов и утрате способности к защите от инфекции. Расстройства регуляции в функционировании данной системы способствуют развитию аутоиммунных болезней, аллергий (на основе пяти типов гиперчувствительности) и росту опухолей. Важность самого существования иммунной системы иллюстрируется появлением в последние 20 лет новой болезни - СПИДа (синдром приобретённого иммунодефицита), при которой наблюдаются все возможные варианты нарушений со стороны иммунной системы.
Антигенявляется макромолекулой, содержащей чужеродную или собственную информацию, которая представляет собой основу для запуска специфического иммунного ответа; с другой стороны, следует иметь в виду, что любой антиген часто служит для исследователей иммунобиологическим маркером. Общее количество различных антигенов оценивается числом 1018. Молекула полного антигена состоит из информаци-
онной части (низкомолекулярные антигенные детерминанты, эпитопы, гаптены) и несущей части (макромолекулярный белок). Изолированная информационная часть сама по себе не является иммуногенной, т.е. не способна индуцировать иммунный ответ. К полным антигенам относятся белки, полисахариды, фосфолипиды.
Антитело,или иммуноглобулин,- это один из типов молекул иммунной системы. Антитела и антигенраспознающие рецепторы могут связывать соответствующие антигены.
Лимфоцитыявляются главными клетками иммунной системы. По существу, иммунная система - это иерархическая совокупность лимфо-идных клеток (1013). Существуют Т-, В- и NK-лимфоциты.Т-клетки дифференцируются в гимусе и играют решающую роль во всех направлениях специфического иммунного ответа. В-лимфоциты дифференцируются в костном мозге, являются предшественниками плазмоцитов - антителопродуцентов. NK-клетки (естественные киллеры) участвуют в неспецифической цитоток-сичности по отношению к внутриклеточно расположенным патогенам. Остальные клетки (макрофаги, нейтрофилы, эозинофилы, дендритные и тучные клетки и др.) также участвуют во многих иммунных процессах, но их участие скорее опосредованное, т.е. они привлекаются лимфоцитами для реализации функций иммунной (лимфоидной) системы.
Стратегической функциейиммунной системы является реализация генетической программы индивидуального развития организма от рождения до смерти в условиях чужеродного окружения. Тактическими функциямииммунной системы являются:
1. Защита от «несвоего» (инфекции, трансплантата).
2. Элиминация модифицированного «своего» (опухолей, повреждённых, стареющих клеток).
3. Регуляция роста и развития клеток и тканей.
Главными партнёрами иммунной системы выступают центральная нервная и эндокринная системы, печень, которые наиболее важны для обеспечения регуляции гомеостаза.
Функциональная организация иммунной си-Часть I. ОБЩАЯ НОЗОЛОГИЯ
стемы может быть рассмотрена на органном, клеточном и молекулярном уровнях. Существуют два типа органов иммунной системы: центральные (или первичные) и периферические (или вторичные). Костный мозг - центральный орган,в котором рождаются все клетки иммунной системы и созревают В-лимфоциты (В-лим-фопоэз, или В-коммитмент). Тимус - центральный орган,в котором дифференцируются Т-лим-фоциты (Т-лимфопоэз, или Т-коммитмент). Кроме того, он является общекоординирующим для всей иммунной системы.
В периферических органах происходит встреча лимфоцитов с антигенами и их специфическая антигензависимая дифференцировка. Этот процесс называется иммунным ответом,сутью которого является создание целой «армии» специфически реагирующих лимфоцитов и специфических антител, которые осуществляют эффек-торные реакции по уничтожению данного конкретного антигена. Существуют Т-клеточный и В-клеточный (гуморальный) пути иммунного ответа.
К периферическим органамотносятся:
1. Лимфатические узлы, лимфатические протоки и селезёнка.
2. Лимфоидная ткань, ассоциированная со слизистыми оболочками (Mucous-Associated Lymphoid Tissue - MALT), которая расположена на пяти уровнях. Первые два уровня - евстахиева труба (Tube-Associated Lymphoid Tissue - TALT) и носоглотка (Nasal-Associated Lymphoid Tissue - NALT) - представлены различными миндалинами; следующий уровень - бронхи (Bronchus-Associated Lymphoid Tissue - BALT) игрудные железы (у женщин); четвёртый уровень - верхние отделы желудочно-кишечного тракта (Gut-Associated Lymphoid Tissue - GALT) - содержат лимфоидную ткань желудка и пейеровы бляшки тонкого кишечника; пятый уровень - это нижние отделы желудочно-кишечного тракта и мочеполовая система - аппендикс, солитарные фолликулы толстого кишечника и лимфоидная ткань мочеполовой системы.
3. Лимфоидная ткань, ассоциированная с кожей (Skin-Associated Lymphoid Tissue - SALT).
Большой вклад в развитие иммунологии от Edward Jenner до наших дней внесли L. Pasteur, И.И. Мечников, P. Ehrlich, Н.Ф. Гамалея, F. Мер. Burnet, N.K. Jerne, P.В. Петров, S. Tonegawa и др.
6.1.2. Клетки иммунной системы
Все клетки, относящиеся к иммунной системе и привлекаемые ею для обеспечения эффек-торных реакций, в функциональном отношении можно разделить начетыре группы:
1. Антигенпрезентирующиеклетки: макрофаги; дендритные клетки типов 1 и 2; В-лимфоциты.
2. Регуляторныеклетки: Т-индукторы; Т-хел-перы типов 1, 2 и 3; Т-регуляторы типа 1.
3. Эффекторные клетки:плазматические клетки (дифференцирующиеся из В-лимфоци-тов); цитотоксические Т-клетки с фенотипом CD8+ (или Т-киллеры); эффекторные Т-клетки воспаления с фенотипом CD4+ (или Т-лимфоци-ты, ответственные за гиперчувствительность замедленного типа); нейтрофилы, эозинофилы, базофилы, тучные клетки, натуральные киллеры (NK-клетки), макрофаги.
4. Клетки памяти:Т-клетки памяти с фенотипом CD8+; Т-клетки памяти с фенотипом CD4+; долгоживующие плазматические клетки; В-клетки памяти.
Номенклатура CD («Cluster of Differentiation») основана на моноклональной технологии,которая была разработана лауреатами Нобелевской премии (1984) G.J.F. КбЫег (Швейцария) и C.Milstein (Аргентина/Великобритания). Она позволяет идентифицировать клетки в зависимости от их происхождения, стадии дифферен-цировки, функционального состояния и т.д. (табл.22). Данная технология, без сомнения, оказалась революционной в иммунологических и смежных областях исследований.
Лимфоциты как главные клетки иммунной системы имеют отличительные особенности:
1. Постоянная «патрульная» рециркуляция по кровотоку, лимфотоку, межтканевым пространствам и секретам.
2. Способность распознавать, т.е. взаимодействовать со «своим» и «несвоим» по принципу «лиганд - рецептор».
3. Клональная организация (F.McF. Burnet) и способность формировать сетевые элементы (N.K. Jerne).
4. Способность к непрерывным реаранжиров-кам в своём геноме в любом возрасте в связи с потребностями формирования специфического ответа на патоген.
5. Умение запоминать о факте встречи с ка-
Глава б / РОЛЬ ИММУННОЙ СИСТЕМЫ В ПАТОЛОГИИ
Таблица 22 Главные идентификационные CD-маркеры клеток иммунной системы
ким-либо антигеном и обеспечивать в будущем экспрессный высокоэффективный ответ на него. Клон - это группа лимфоцитов, коммитиро-ванная к определённому антигену. До встречи с этим антигеном каждый лимфоцит клона называют наивным. По-видимому, в человеческом организме исходно существуют десятки миллионов клонов Т- и В-лимфоцитов. После встречи с соответствующим антигеном и в результате иммунного ответа коммитированный лимфоцит становится праймированным.
6.1.3. Молекулы иммунной системы
Для осуществления необходимых функций клетки иммунной системы имеют сложную молекулярную организацию своих рецепторов и способны к выработке целого ряда молекул.
1. Антигенпредставляющие, антигенраспоз-
нающие и антигенсвязывающие молекулы.
Набор этих молекул уникален для каждого орга
низма, для каждого лимфоцитарного клона и для
каждого специфического иммунного ответа. К
ним относятся:
- антигенраспознающие иммуноглобулиновые рецепторы В-клеток (В cellular receptors - BCR);
- свободные иммуноглобулины: IgM, IgG, IgA, IgE, IgD;
- антигенраспознающие рецепторы Т-клеток (T cellular receptors - TCR);
- антигенпредставляющие молекулы: лейкоцитарные антигены главного комплекса гисто-совместимости (Human leukocyte antigens -HLA I и HLA II) и молекулы CD1 (a, b, c, d, e).
2. Адгезивные молекулы опосредуют взаи
модействия между клетками и лигандами при
непосредственном контакте:
- суперсемейство иммуноглобулинподобных молекул;
- интегрины;
-
-
- селектины;
- муцины;
- суперсемейство рецепторов к факторам некроза опухоли и фактору роста нервов;
- линк-семейство (компоненты экстрацеллю-лярного матрикса).
3. Иммуноцитокины являются гормонами
иммунной системы, действующими чаще с пара-
и аутокринным, реже - с эндокринным эффекта
ми:
- интерлейкины (ИЛ);
- колониестимулирующие факторы (КСФ);
- интерфероны (ИФН);
- факторы некроза опухоли (ФИО);
- хемокины и др.
4. Сборная группа различных медиаторов
иммунного воспаления включает белки комп
лемента, «острой фазы», простаноиды и лейко-
триены, протеолитические ферменты и др.
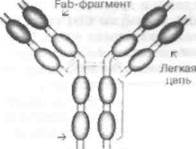
Иммуноглобулины М, G, А, Е и D - это эф-фекторные молекулы гуморального иммунного ответа. Молекула иммуноглобулина является гликопротеином; белковые цепи включают сотни аминокислотных последовательностей; углеводный компонент составляет 12 %. Молекула IgG (рис. 36) состоит из двух идентичных лёгких (light - L) и двух идентичных тяжёлых (heavy - Н) цепей. Лёгкие цепи бывают двух типов: х и X, а тяжёлые - пяти (ц, у, а, е и 8). Лёгкие и тяжёлые цепи содержат повторяющиеся гомологичные последовательности и образуют своеобразные «клубки» (домены). Существуют константные (constant) (CL, CHI, CH2 и СНЗ) и вариабельные (variable) домены (VL и VH). Гипервариабельные области вариабельных доменов формируют антигенсвязывающие сайты, или активные центры. Молекула иммуноглобулина может быть также разделена на Fc-фрагмент (а fragment crystalline), который ответствен за не-
Часть I. ОБЩАЯ НОЗОЛОГИЯ
специфическую эффекторную активность, и 2 Fab-фрагмента (fragment antigen-binding), которые содержат антигенсвязывающие сайты. Химическая природа антител была подробно изучена лауреатами Нобелевской премии (1972) G.M. Edelman (США) и R.R. Porter (Великобритания).
Каждый В-лимфоцит экспрессирует В-клеточ-ный антигенраспознающий рецептор(BCR), который состоит из мономерных иммуноглобулинов М и D, имеет клональную гетерогенность и ассоциирован с молекулами CD79a и CD79P, необходимыми для проведения сигнала внутрь клетки. Наряду с этими молекулами имеется также корецепторный комплекс (CD19, CD21(CR2), CD81), предназначенный для распознавания HLA П.
Каждый Т-лимфоцит экспрессирует Т-кле-точный антигенраспознающий рецептор(аР TCR), который состоит из двух цепей (а и Р) и ассоциирован с одним из корецепторов - CD4 (у Т-хелперов) или CD8 (у цитотоксических Т-лим-фоцитов). Эти инвариантные корецепторы CD4 и CD8 необходимы для распознавания соответственно HLA II или HLA I. Каждая цепь TCR подобно молекуле иммуноглобулина имеет вариабельный и константный домены, что обеспечивает клональную гетерогенность антигенраспоз-нающих рецепторов. Ещё одна молекула (CD3) тесно ассоциирована с TCR и служит для проведения сигнала внутрь клетки. CD3 состоит из 5 инвариантных белков (е,у,5,^ч)- Другой тип ан-тигенраспознающего рецептора - y5TCR - эксп-рессируется на небольшой субпопуляции Т-кле-ток. Эти у5Т-клетки, роль которых пока до конца не исследована, по-видимому, функционируют подобно NK-клеткам, однако они тоже имеют определённую клональную гетерогенность.
Молекулы гистосовместимостибыли открыты лауреатами Нобелевской премии (1980) В. Benacerraf (США), J. Dausset (Франция) и G.D. Snell (США). Эти молекулы играют решающую роль во многих иммунных процессах, включая загрузку антигенных пептидов и их презентацию. Молекулы HLA разделяются на класс I (А, В, С, Е, F, G) и класс II (DR, DP, DQ) взависимости от их структуры и функции. Экспрессия HLA I имеет место почти на всех клетках (за исключением синцитиотрофобласта), выполняя функцию взаимной информации клеток внутри организма об аутологичности; экспрессия HLA II наблюдается почти исключительно на клет-
ках иммунной системы: В-лимфоцитах, макрофагах, эндотелиоцитах, активированных Т-клет-ках и др.
| Fc-фрагмент |
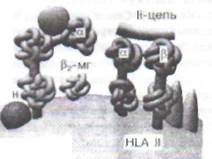
 Молекула HLA I(рис.37) состоит из трехдо-менной сс-цепи и Р2-микроглобулина, связь между которыми стабилизируется калнексином. Между доменами а, и а2 имеется желобок,предназначенный для нагрузки антигена. Моею того как антиген загружается, происходит высвобождение Р2-микроглобулина, возможно, за счёт кон-формационных изменений. Имеется около 180 генов с более чем 500 аллелями (на хромосоме
Молекула HLA I(рис.37) состоит из трехдо-менной сс-цепи и Р2-микроглобулина, связь между которыми стабилизируется калнексином. Между доменами а, и а2 имеется желобок,предназначенный для нагрузки антигена. Моею того как антиген загружается, происходит высвобождение Р2-микроглобулина, возможно, за счёт кон-формационных изменений. Имеется около 180 генов с более чем 500 аллелями (на хромосоме
Тяжелая цепь
Рис. 36. Структура молекулы IgG. Молекула IgG состоит их двух идентичных легких и двух идентичных тяжелых цепей, которые образуют три фрагмента: два Fab (антисвязывающие сайты) и один Fc (неспецифические эффекторные функции). Цепи укладываются таким образом, что возникают «клубки» - домены, которые могут быть вариабельными или константными
| Тапази |
| HLA I |
 Кальретикулин
Кальретикулин
Рис. 37. Структура белков гистосовместимости (HLA I и HLA II) и шапероны. Молекула HLA I состоит из а-цепи (три домена) и Р2-микроглобулина (Р2-мг). Молекула HLA II содержит две цепи: а (два домена) и р (тоже два домена). Шапероны (кальретикулин, тапазин, Ii-цепь и др.) - это белки, ответственные за правильную укладку белковых молекул; в частности, шапероны обеспечивают протекцию «желобков» молекул HLA до момента загрузки на них антигенных пептидов
Глава б / РОЛЬ ИММУННОЙ СИСТЕМЫ В ПАТОЛОГИИ
б), кодирующих синтез молекул HLA I, что обеспечивает их видоспецифический ииндивидуальный полиморфизм. Молекула HLA IIсостоит из двух димеров - а и р. Вариабельные домены -а, ир, - формируют желобок для загрузки антигена. В целом Р-цепи HLA-DR, DP, DQ характеризуются меньшей гетерогенностью; известно более 300 аллельных продуктов HLA II (более 20 генов, кодирующих HLA II, также находятся на хромосоме 6). Для контроля укладки при синтезе, протекции, транспорте и загрузке антигенных пептидов на HLA служат особые молекулы -шапероны(калнексин, кальретикулин, инвариантная цепь И идр.).
He-HLA молекулы CD1 (а, Ь,с, d,e), которые состоят по аналогии с HLA I из сс-цепи и Р2-микроглобулина, также вовлечены в процессы загрузки антигенов, но небелковой природы (фос-фолипиды, липополисахариды).
Имеется связь между наследованием некоторых генов HLA и высоким риском развития определённых болезней. Например, более 90% пациентов с анкилозирующим спондилоартритом, тяжёлой аутоиммунной патологией позвоночника имеют ген HLA-B27.
Поскольку HLA определяет гистосовмести-мость, необходимо, чтобы у донора и реципиента при трансплантации органа или ткани было соответствие по HLA. Карта пациента по HLA называется «full house» («полный дом»)и включает данные относительно двух аллелей каждого вида молекул (например, HLA-A, HLA-B, HLA-DR и др.). Сенсибилизация белками HLA в прошлом (при гемотрансфузиях, трансплантациях или беременности) может приводить к острому отторжению пересаживаемого трансплантата или тромбоцитопении при переливании крови, поэтому необходимо обязательное тестирование на наличие антител против HLA. Типирование HLA может служить дополнительным критерием для диагностики таких болезней, как анки-лозирующий спондилоартрит, диабет, целиакия, гемохроматоз, псориаз и нарколепсия, при которых известна высокая степень связи с определёнными гаплотипами HLA.
6.2. ИММУННЫЙ ОТВЕТ
6.2.1. Стадии иммунного ответа
Иммунный ответ- это процесс взаимодей-
ствия клеток иммунной системы, который индуцируется антигеном и приводит к образованию эффекторных клеток и молекул, уничтожающих данный антиген. Иммунный ответ является всегда специфическим,но не изолированным процессом, который протекает только в периферических органах иммунной системы. Как правило, он сопровождается такими неспецифическими реакциями,как фагоцитоз, активация комплемента, NK-клеток ит.д.
В начальных стадиях иммунного ответа участвуют по крайней мере три вида клеток: макрофаг (а также дендритная клетка), Т- и В-лим-фоциты. В целом все клетки, вовлечённые в этот процесс, могут быть разделены, как указывалось выше, на антигенпредставляющие, регулятор-ные, эффекторные и клетки памяти. Имеются два магистральных пути иммунного ответа:
Дата добавления: 2015-07-14; просмотров: 955;