Деградация Н2О2.
Как и большинство АФК, пероксид водородатоксичен для клеток, и
поэтому его концентрация in vivoстрого регулируется. Множество ферментативных и неферментативных антиоксидантных систем внутри клетки защищают ее от токсичного действия Н2О2. Многие ферментативные антиоксидантные системы также принимают участие в сигнальных путях, контролируя и регулируя передачу сигнала. Основные белки, участвующие
в деградации Н2О2 –это каталазы, пероксиредоксины и глутатионпероксидазы.
Каталазы –это гемсодержащие ферменты, которые разлагают Н2О2непосредственно до воды и кислорода (уравнение 10):
2H2O2 - 2H2O + O2(10)
Каталазы есть у большинства аэробных клеток. Эти ферменты вносят значительный вклад в деградацию Н2О2, защищая организм от окислительного стресса, а также ограничивают участие Н2О2в сигналинге. В эукариотических клетках каталазы локализуются в пероксисомах и считаются ключевыми ферментами, осуществляющими элиминацию «нежелательных» АФК. Величина Кm каталазы по пероксиду водорода имеет миллимолярную размерность. Поэтому этот фермент эффективен при деградации высоких концентраций оксиданта.
Пероксиредоксины (далее Prx) – это семейство белков, катализирующих восстановление Н2О2 и алкилгидропероксидов до воды и спирта. Для этого процесса пероксиредоксины используют восстанавливающие эквиваленты, предоставляемые тиоредоксином, тиоредоксинредуктазой (TR) и NADPH(уравнение 11):
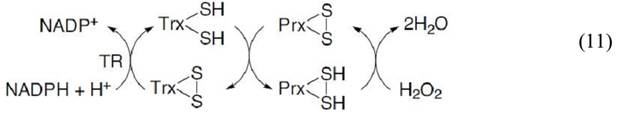
По строению активного центра выделяют три группы пероксиредоксинов: типичные 2Cys-Prx, атипичные 2Cys-Prx и 1Cys-Prx. Выделяемые таким образом группы различаются количеством и консервативностью расположения цистеиновых остатков. У млекопитающих, как и у большинства эукариот, представители типичных 2Cys-Prx взаимодействуют с Н2О2 с образованием сульфеновой кислоты, которая затем быстро преобразуется в внутрицепочечный дисульфид. Иногда в присутствии Н2О2 сульфеновая кислота взаимодействует со второй молекулой Н2О2 и окисляется до сульфиновой кислоты, что таким образом инактивирует фермент. Участие пероксиредоксинов в регуляции концентрации Н2О2 позволяет предположить, что переокисление может служить важным механизмом, благодаря которому в некоторых случаях Н2О2 имеет возможность избежать деградации и принять участие в сигнальных каскадах. В дальнейшем, роль этого механизма в редокс-сигналинге была подтверждена благодаря открытию сульфиредоксинов и сестринов, способных восстанавливать сульфиновую кислоту пероксиредоксинов до тиола. Кроме того, экспрессия генов этих белков
регулируется в ответ на окислительный стресс. Пероксиредоксины локализуются почти во всех компартментах эукариотических клеток. Физиологический диапазон концентраций пероксида водорода, с которыми способны справиться пероксиредоксины составляет 1-10 мкM. Превышение этих концентраций вызывает переокисление и инактивацию пероксиредоксинов, что может иметь и защитную функцию. Дело в том, что восстановление пероксиредоксинов требует постоянного расхода NADPH. При окислительном стрессе пероксиредоксины становятся основным субстратом для тиоредоксина, быстро истощая пул NADPH. Переокисленный пероксиредоксин перестает быть субстратом тиоредоксина. Клетка перестает тратить NADPH на борьбу с оксидантом, сохраняя ценный субстрат для восстановления других белков, так в борьбе с пероксидом водорода подключается каталаза, имеющая высокое сродство к пероксиду и использующая одну молекулу Н2О2 для восстановления другой.
Глутатионпероксидазы, которые совмещают процесс восстановления Н2О2 до воды с окислением глутатиона до глутатиондисульфида (уравнение 12):
H2O2+ 2GSH - GSSG + 2H2O (12)
Это пероксидазы, многие из которых содержат в активном центре остаток
селеноцистеина, составляют группу, имеющую как минимум четыре изоформы. Глутатионпероксидазы широко представлены в тканях животных, а также в некоторых водорослях и грибах, но отсутствуют у высших растений и бактерий. Глутатионпероксидазы специфичны к глутатиону как к электрон-донору и могут взаимодействовать с другими пероксидами. Как представители ферментов, деградирующих Н2О2, глутатионпероксидазы
контролируют сигналинг, опосредованный Н2О2, а также могут осуществлять редокс-сигналинг путем окисления глутатиона, который далее модифицирует белки через реакцию тиол-дисульфидного обмена.
Дата добавления: 2015-08-11; просмотров: 1681;