НУКЛЕИНОВЫЕ КИСЛОТЫ И НУКЛЕОПРОТЕИДЫ
Нуклеиновые кислоты – высокомолекулярные гетерополимеры, играющие основную роль в сохранении и реализации генетической информации. Различают два типа нуклеиновых кислот (НК) – дезоксирибонуклеиновые кислоты ДНК, которые обеспечивают сохранение информации, и рибонуклеиновые кислоты РНК, принимающие участие в процессах генной экспрессии и биосинтеза белка. Нуклеиновые кислоты построены из нуклеотидных звеньев, которые в свою очередь состоят из азотистого основания, углеводного остатка и фосфатной группы.
При мягком щелочном гидролизе нуклеиновые кислоты распадаются на мономеры – нуклеотиды. Нуклеотиды при нагревании до 145оС с водным аммиаком теряют остаток фосфорной кислоты с образованием нуклеозидов. Нуклеозиды в условиях кислотного гидролиза распадаются на азотистые основания и сахара.
Азотистые основания – это ароматические гетероциклические соединения, производные пиримидина или пурина. Пять азотистых оснований являются основными структурными компонентами нуклеиновых кислот, общими для всей живой материи.
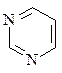
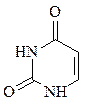
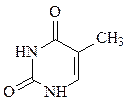
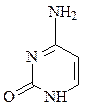
пиримидин урацил тимин цитозин
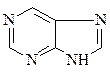
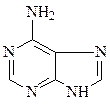
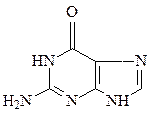
пурин аденин гуанин
В состав ДНК входят аденин, гуанин, тимин и цитозин. В состав РНК входят аденин, гуанин, урацил и цитозин. Кроме перечисленных азотистых оснований, в нуклеиновых кислотах в небольших количествах встречаются модифицированные азотистые основания, в основном гидроксилированные и метилированные, их называют минорными.
Для пуриновых и пиримидиновых оснований характерны лактам-лактимная и амин-иминная таутомерия. По данным ИК- и ЯМР-спектро-скопии выявлено, что в нуклеиновых кислотах пуриновые и пиримидиновые основания преимущественно находятся в лактамной и аминной формах, это обеспечивает правильность спаривания.
Азотистые основания поглощают свет в УФ области спектра с макси-мумом 260 нм, что используется для количественного определения НК.
Углеводная часть нуклеотидов, входящих в РНК, представлена рибозой, а входящих в ДНК, – дезоксирибозой. Пентозы в нуклеиновых кислотах всегда присутствуют в β-D-фуранозной форме. Углеводные атомы пентоз в нуклеотидах нумеруют со знаком «штрих», чтобы их можно было отличить от атомов азотистых оснований.
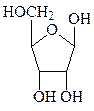
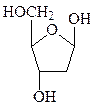
рибоза дезоксирибоза
Соединения азотистых оснований с пентозой с помощью N-гликозид-ных связей называют нуклеозидами – рибонуклеозидами (аденозин, гуанозин, цитидин и уридин) и дезоксирибонуклеозидами (дезоксиаденозин, дезоксигуанозин, дезоксицитидин и тимидин).
Нуклеозиды, этерифицированные фосфорной кислотой по 5’-положе-нию, называются нуклеотидами. В случае одного остатка фосфорной кислоты они называются нуклеотидмонофосфатами, в случае двух остатков – нуклеотиддифосфатами, в случае трех – нуклеотидтрифосфатами.
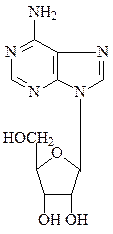 аденозин
аденозин 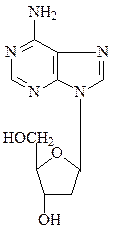 дезоксиаденозин
дезоксиаденозин
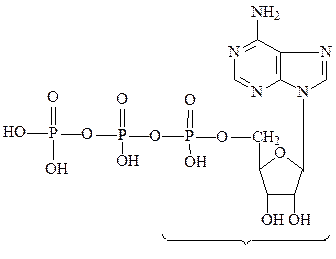 аденозин-5’-монофосфат (АМФ)
аденозин-5’-монофосфат (АМФ)
 аденозин-5’-дифосфат (АДФ)
аденозин-5’-дифосфат (АДФ)
 аденозин-5’-трифосфат (АТФ)
аденозин-5’-трифосфат (АТФ)
В молекулах нуклеотидтрифосфатов остатки фосфорной кислоты соединены ангидридной связью, обладающей большой потенциальной энергией. Такие связи называются макроэргическими, а соединения – высокоэнергетическими. АТФ занимает центральную роль в обмене веществ и энергией во всех клетках, ГТФ энергетически обеспечивает процессы трансляции белков, УТФ необходим для синтеза гликогена, ЦТФ участвует в синтезе глицерофосфолипидов.
Нуклеиновые кислоты – это неразветвленные полимеры нуклеотидов, соединенных между собой 3’,5’-фосфодиэфирной связью. Полинуклеотиды, составленные из рибонуклеотидных звеньев, называются рибонуклеиновыми кислотами (РНК), из дезоксирибонуклеотидных звеньев – дезоксирибонуклеиновыми кислотами (ДНК).
Ионизированные фосфатные группы полимерного остова сообщают молекулам нуклеиновых кислот большой отрицательный заряд. По этой причине в клетках ДНК обычно присутствуют в комплексе: 1) с основными белками – гистонами и протаминами; 2) с поликатионами типа спермидина H2N-(CH2)4-NH-(CH2)3-NH2; 3) с щелочноземельными катионами – Mg2+.
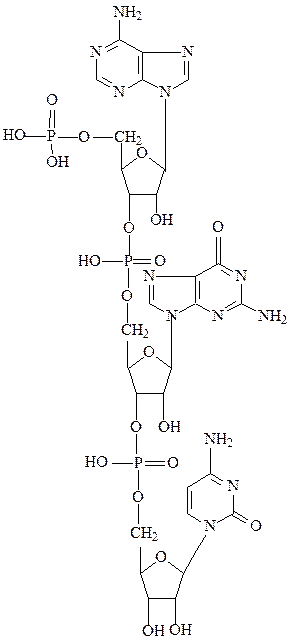
пример нуклеотида, построенного по типу РНК:
В РНК во 2-ом положении рибозы имеются свободные гидроксильные группы, сильно влияющие на конформацию полимера. Отсутствие 2-ОН-группы в дезоксирибозе делает ДНК более устойчивой к щелочной деградации.
Как ДНК, так и РНК легко гидролизуются под действием кислот. Гликозидные связи в ДНК более лабильны, чем в РНК. Пуриновые нуклеотиды гидролизуются легче, чем пиримидиновые, и все же ковалентная структура этих молекул сравнительно устойчива, что определяет их способ-ность служить генетическим материалом. Однако как ДНК, так и РНК вступают во множество химических реакций по азотистым основаниям:
а) азотистая кислота реагирует с аминогруппами оснований, превращая их в гидроксильные группы, которые подвергаются таутомеризации и превраща-ются в кетогруппы. Таким образом, азотистая кислота превращает цитозин в урацил, аденин – в гипоксантин, гуанин – к ксантин и, следовательно, является эффективным химическим мутагеном.
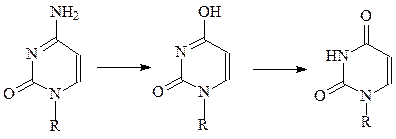
цитозин урацил
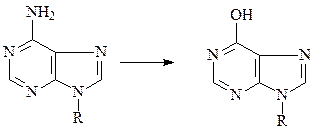
аденин гипоксантин
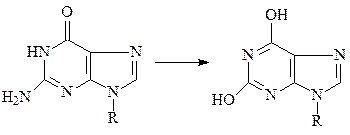
гуанин ксантин
б) другое мутагенное соединение, гидроксиламин (H2N-OH), легко реагирует с карбонильными группами, особенно в пиримидинах, даже несмотря на то, что эти карбонильные группы являются частью циклической амидной структуры и поэтому имеют сравнительно низкую реакционную способность.
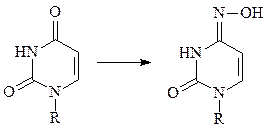
в) пиримидины легко восстанавливаются боргидридом натрия NaBH4.

г) уридин, тимидин и, в меньшей степени, цитозин в составе нуклеиновых кислот димеризуются при УФ-облучении; цитозин под действием УФ-света легко гидратируется.
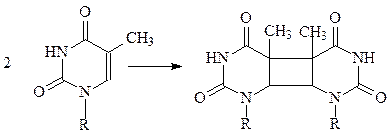
тимин
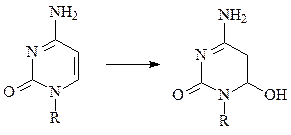
цитозин
д) формальдегид вступает в реакцию с аминогруппами пуриновых и пирими-диновых оснований.
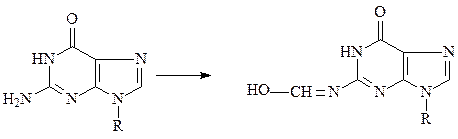
гуанин
Биологической ролью ДНК является хранение информации о белках, необходимых для жизни клетки, и передача этой информации дочерним клеткам.
Структурная организация ДНК достаточно сложна и имеет три уровня.
Первичная структура ДНК – это линейная цепь четырех видов дезоксирибонуклеотидов, соединенных фосфодиэфирной связью.
Вторичная структура ДНК – это двухцепочечная правозакрученная спираль из комплементарных друг другу антипараллельных полинуклеотид-ных нитей. Модель была предложена в 1953 г Уотсоном и Криком.
Свойство комплементарности было выведено на основании некоторых закономерностей, отмеченных Чаргаффом, а именно:
- в каждом типе ДНК количество аденина примерно равно количеству тимина, а количество гуанина – количеству цитозина;
- сумма пуриновых оснований равна сумме пиримидиновых;
- напротив, соотношение (аденин+тимин)/(гуанин+цитозин) в различных организмах неодинаково.
Было установлено, что АТ-тип ДНК преобладает в животных и расти-тельных клетках, а ГЦ-тип – у микроорганизмов. Сравнение организмов по их ГЦ-содержанию иногда используют как основу для установления филогенетического родства. Нужно, однако, учесть, что тимин под действием УФ-света претерпевает ряд фотохимических превращений, поэтому бактерии с высоким ГЦ-содержанием могут чаще встречаться там, где много солнечного света, тогда как более низкое ГЦ-содержание может указывать на то, что местообитанием данного организма являются более защищенные от солнца места.
Модель Уотсона и Крика позволила объяснить причину этих закономерностей – каждое основание одной цепи связано с комплементар-ным ему основанием другой цепи водородными мостиками. При этом аденин комплементарен тимину, а гуанин – цитозину. Таким образом, каждая пара состоит из одного пуринового и одного пиримидинового основания, причем пары комплементарных оснований близки и по размеру, и по форме. Пара А-Т может образовывать два, а пара Г-Ц даже три водородных мостика.
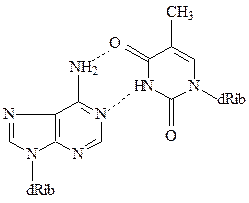
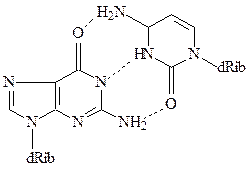
Благодаря способности азотистых оснований к спариванию, образуется хорошо стабилизированная двухцепочечная структура, где пары азотистых оснований расположены внутри двойной спирали и образуют неполярную область ДНК. Гидрофобные взаимодействия между ароматическими азотистыми основаниями стабилизируют структуру, преодолевая силы электростатического отталкивания между отрицательно заряженными фосфатными группами. Остатки дезоксирибозы двух антипараллельных цепей образуют правозакрученную спираль, где на каждый виток приходится 10 пар оснований. Внешняя сторона спирали заряжена отрицательно за счет фосфатных групп. В спирали ДНК существуют две бороздки – большая и маленькая.
На основании рентгеноструктурного анализа было установлено, что двойная спираль ДНК кроме описанной выше формы В, благоприятной для репликации, может существовать в некоторых других формах: форме А, благоприятной для процессов транскрипции; форме Z, участвующей в регуляции экспрессии генов; форме С, выявленной у ряда вирусов. Все эти формы различаются диаметром, числом пар оснований в витке, углом наклона плоскости оснований по отношению к оси молекулы и способны взаимно переходить друг в друга в зависимости от условий среды.
В функциональном отношении две цепи ДНК не эквивалентны. Кодирующей цепью является та из них, которая считывается в процессе транскрипции. Именно эта цепь служит матрицей для РНК. Некодирующая цепь по последовательности подобна РНК при условии замены тимина на урацил. Общепринято давать структуру гена в виде последовательности некодирующей цепи ДНК в направлении 5’→3’. Если прочитать кодоны в этом направлении, то с помощью генетического кода можно воспроизвести аминокислотную последовательность белка в принятом порядке, от N-конца к С-концу.
Третичная структура ДНК – более плотная упаковка с образованием сложных трехмерных структур. Прокариоты имеют кольцевую ДНК, закрученную в левую суперспираль. Эукариоты также образуют суперспираль, но не свободной ДНК, а ее комплексов с белками хромосом.
Дезоксирибонуклеопротеиды – комплексы ДНК с белками хромосом – составляют хроматин. Хроматин можно видеть в оптический микроскоп только во время деления клеток, когда он находится в конденсированном виде в составе хромосом. Гистоны формируют первичную степень компактизации хроматина – так называемые нуклеосомы. По две молекулы каждого из гистонов Н2а, Н2б, Н3, Н4 составляют октамер (кор), обвитый 1,8 витками спирали ДНК поверх белковой структуры. Участок ДНК (линкерная ДНК), непосредственно не связанная с гистоновым кором, взаимодействует с гистоном Н1. Этот белок обеспечивает формирование вторичной суперспиральной структуры ДНК – нуклеомеры. Когда хроматин конденсируется далее за счет тех же Н1 как скрепок формируются петли – так называемая структура «ламповых щеток». Примерно 20 петель образуют минидиски – хромонемы. Большое число минидисков укладывается в стопку, составляя хромосому. Гистоны в октамере имеют подвижный N-конец из 20 аминокислот, который выступает из нуклеосомы и важен для поддержания структуры хроматина и контроля за генной экспрессией. Так, конденсация хромосом связана с фосфорилированием гистонов, а усиление транскрипции – с ацилированием в них остатков лизина. В хроматин входит также группа негистоновых белков. Она высоко гетерогенна и включает структурные ядерные белки, множество ферментов и факторов транскрипции, связанных с отдельными участками ДНК и осуществляющих регуляцию генной экспрессии и других процессов.
РНК принимают участие во всех стадиях процесса генной экспрессии и биосинтеза белка. Различают три основных вида РНК: рибосомальная (рРНК), транспортная (тРНК) и матричная (мРНК), которую также называют информационной (иРНК). РНК клетки существенно различаются по размерам и продолжительности существования, по вторичной и третичной структуре. Из-за стерических ограничений, вызванных ОН-группой во 2’-положении рибозы, РНК не могут образовывать структур, подобных двойной спирали и имеют менее регулярную вторичную структуру.
мРНК (иРНК) – короткоживущие и наиболее гетерогенные РНК. Они являются копиями определенных участков ДНК, в которых заключена информация о последовательности аминокислот в кодируемых ими белках. После синтеза мРНК транспортируются к рибосомам – белоксинтезирующим машинам клетки, где на их основе синтезируется необходимый клетке белок. В целом, в линейной молекуле мРНК формируется несколько двухспиральных шпилек, на концах которых располагаются знаки инициации и терминации трансляции.
тРНК, активирующие аминокислоты и переносящие их к рибосомам, являются небольшими долгоживущими молекулами. Они содержат 70–90 нуклеотидов, которые с помощью своих антикодонов узнают за счет спаривания оснований определенные кодоны на мРНК. На 3’-конце всех тРНК, называемом акцепторным участком, обязательным является тринуклеотид ЦЦА, к которому присоединяется определенная аминокислота. Каждой из 20 аминокислот соответствует своя тРНК. Для некоторых аминокислот, кодируемых двумя или более кодонами, существует несколько тРНК. В молекуле тРНК содержится довольно много минорных и модифи-цированных оснований – псевдоуридин, дигидроуридин, тимидин, встречающийся обычно в ДНК, а также много метилированных нуклеотидов, таких как 7-метилгуанидин. Конформацию молекулы стабилизируют многочисленные пары оснований, часть из которых не соответствует общим принципам спаривания оснований. Пространственное изображение вторичных структур всех тРНК имеет форму клеверного листа. В «клеверном листе» кроме акцепторного участка различают три обязательные петли:
а) антикодоновая петля – содержит антикодон, представляющий собой специфический триплет нуклеотидов, который комплементарен в анти-параллельном направлении кодону мРНК, кодирующему соответствующую аминокислоту;
б) псевдоуридиловая петля – обеспечивает взаимодействие с рибосомами;
в) дигидроуридиловая петля – обеспечивает взаимодействие тРНК с соот-ветствующей аминоацил-тРНК-синтетазой.
Третичная структура тРНК очень компактна и чаще всего описывается термином «локтевой сгиб». Третичные структуры всех тРНК настолько похожи, что смесь различных тРНК образует кристаллы.
Самой длинной и долгоживущей является рРНК, выполняющая роль структурного компонента рибосом. Число рибосом в клетке очень велико. Вторичная структура рРНК образуется за счет коротких двухспиральных участков молекулы – шпилек, чередующихся с одноцепочечными участками, к которым присоединяются белки. Третичная структура рРНК – образование в комплексе с рибосомальными белками малой и большой субъединицы рибосом.
Нуклеиновые кислоты могут денатурировать. Этот процесс состоит в расхождении цепей двойной спирали ДНК и двухспиральных участков молекулы РНК. Денатурацию можно вызвать нагреванием, добавлением кислоты, щелочи, спиртов, мочевины или удалением стабилизирующих структуру молекулы ДНК противоионов, например Mg2+. В результате денатурации каждая из цепей молекулы приобретает форму беспорядочно свернутого клубка.
Денатурация ДНК сопровождается усилением оптического поглощения в УФ-области пуриновых и пиримидиновых оснований. Это явление называют гиперхромным эффектом. В нативном состоянии нуклеиновые кислоты поглощают свет менее интенсивно, чем в денатурированном. Это явление называют гипохромным эффектом. Гипохромный эффект обусловлен гидрофобными взаимодействиями между основаниями, плотно уложенными в стопку в структуре нативной молекулы. Тепловая денатурация нуклеиновых кислот происходит в довольно узком интервале температур. Характерным параметром процесса является температура плавления. Температурная зависимость поглощения УФ-света при 260 нм называется кривой плавления. Температура плавления (Тпл) – это точка, при которой прирост поглощения составляет половину максимального. Чем выше ГЦ-содержание нуклеиновой кислоты, тем более она становится устойчивой к денатурации, причем зависимость Тпл от ГЦ-содержания почти линейна. Точное соотношение между ГЦ-содержанием ДНК и Тпл очень сильно зависит от ионного состава и рН среды. Полная денатурация молекулы ДНК приводит к расхождению комплементарных цепей. При быстром охлаждении раствора денатурированной ДНК цепи остаются в разделенном состоянии. Однако если в течение какого-то времени поддерживать температуру немного ниже температуры плавления (этот процесс называют отжигом), может вновь восстановиться нативная структура. Этот факт дает в руки ученым очень важный метод исследования нуклеиновых кислот, основанный на образовании гибридов.
Одним из типов двойных спиралей, которые получают искусственным путем, является гибрид ДНК-РНК. Оказалось, что молекулы мРНК гибридизируются с одной из двух разделившихся цепей ДНК, несущей участки, комплементарные мРНК. Метод гибридизации используется также для получения гетеродуплексов ДНК, в которых две цепи молекулы происходят от двух разных генетических линий одного и того же вида организмов. Известно, что некоторые мутации состоят в выпадении или вставке одного или нескольких оснований в цепь ДНК. Гетеродуплексы, в которых одна из цепей нативная, а другая – со вставкой или делецией, имеют по всей своей длине нормальную структуру по Уотсону-Крику, за исключением тех участков, где делеции или вставки нарушают комплементарность и образуются одноцепочечные петли.
Опыты по гибридизации позволили исследовать гомологичность нуклеиновых кислот из разных источников. Вначале молекулы ДНК расщепляют с помощью ультразвука на фрагменты длиной около 1000 нуклеотидов и подвергают денатурации. Фрагменты денатурированной ДНК смешивают с денатурированной ДНК из другого источника. Участки ДНК разных видов, имеющие близкие нуклеотидные последовательности, в той или иной степени гибридизуются, тогда как участки с сильно различающимися последовательностями гибридизации не поддаются.
ЛЕКЦИЯ 9
Дата добавления: 2015-08-08; просмотров: 2778;