УГЛЕВОДЫ
Углеводы – группа природных полигидроксиальдегидов и полигид-роксикетонов с общей формулой (СН2О)n.
В биосфере на долю углеводов приходится больше, чем на все другие органические соединения вместе взятые.
Биологические функции углеводов:
1) источник углерода для синтеза белков, нуклеиновых кислот, липидов и других соединений;
2) источник энергии, покрывают до 70% потребности организма, важные (хотя и не жизненно необходимые) компоненты питания;
3) резервная функция, запасаются в виде полисахаридов;
4) структурная функция, являются строительным материалом для многих организмов;
5) защитная функция, защищают клетки и ткани от высыхания, повреждения чужеродными агентами;
6) участие в образовании гибридных молекул гликопротеинов и глико-липидов.
Различают три основных класса углеводов – моносахариды, дисахари-ды (олигосахариды) и полисахариды.
Моносахариды – простые углеводы, содержат только одну структурную единицу (СН2О)n, где n≥3. В зависимости от числа атомов углерода в молекуле среди моносахаридов различают триозы, тетрозы, пентозы, гексозы и гептозы. В природе наиболее часто встречаются пентозы и гексозы.
В основу номенклатуры положены тривиальные названия преимущественно с окончанием –оза. В зависимости от того является моносахарид альдегидом или кетоном, он будет называться альдозой или кетозой. Альдозы встречаются чаще, чем кетозы. Практически все моносахариды являются D-изомерами (расположение ОН-группы у послед-него хирального атома углерода аналогично конфигурации D-глицеринового альдегида).
Формулы основных моносахаридов в проекции Фишера:
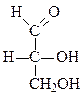
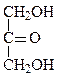
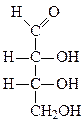
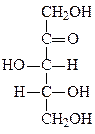
D-глицеральдегид дигидроксиацетон D-эритроза D-ксилулоза
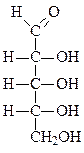

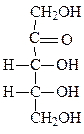
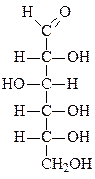
D-рибоза 2-дезокси-D-рибоза D-рибулоза D-глюкоза
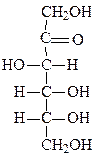
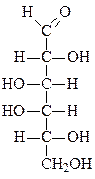
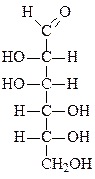
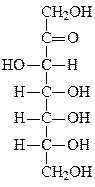
D-фруктоза D-галактоза D-манноза D-седогептулоза
Два сахара, различающиеся по конфигурации вокруг только одного атома углерода, представляют собой эпимеры по отношению друг к другу (глюкоза и манноза по второму атому, глюкоза и галактоза – по четвертому).
В растворах моносахариды из пяти и более атомов углерода существуют в виде замкнутых циклических структур – циклических полуацеталей, образованных в результате взаимодействия карбонильной группы с одной из гидроксильных групп. Сахара с шестичленным циклом называются пиранозами, сахара с пятичленным циклом называются фуранозами. Естественная тенденция пентоз и гексоз к циклизации обеспечивает образование устойчивых колец из чрезвычайно реакционноспособных неустойчивых мономеров.
Циклические формы моносахаридов изображают в проекции Хеуорса: ОН-группы, которые в фишеровской проекции располагаются справа, в про-екции Хеуорса располагаются под плоскостью кольца, а группы, находящиеся слева, – над плоскостью кольца.
Изомерные формы моносахаридов, отличающиеся друг от друга только конфигурацией полуацетального углеродного атома, такие как α-D-глюкоза и β-D-глюкоза, называются аномерами.
Реакции моносахаридов (на примере глюкозы):
1. Мутаротация – переход аномеров из одной формы в другую.
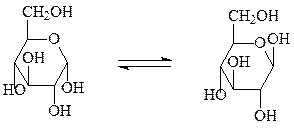
α-D-глюкопираноза (≈40%) β-D-глюкопираноза (≈60%)
2. Окисление – образование лактонов и кислот (по первому и шестому атому углерода).
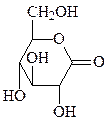
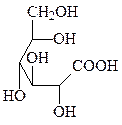
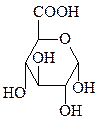
глюконолактон глюконовая кислота глюкуроновая кислота
3. Восстановление – образование сахароспиртов.
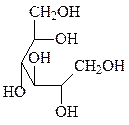 сорбит
сорбит
3. Эпимеризация – в слабощелочном растворе глюкоза находится в равно-весии с фруктозой и маннозой.
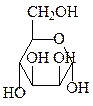
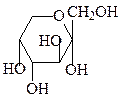
α-D-манноза D-фруктоза
4. Образование гликозидов – конденсация аномерной ОН-группы со спир-товой группой (О-гликозиды) или аминной группой (N-гликозиды). Гликозидная связь – основная связь, с помощью которой образуются полисахариды. Гликозидами также является большое количество низкомолекулярных регуляторов у растений. Неуглеводная часть молекулы гликозида называется агликоном.
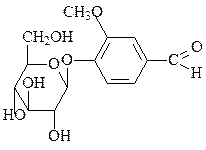
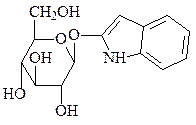
ванилин-α-D-глюкозид (источник ванили) индиган (источник индиго)
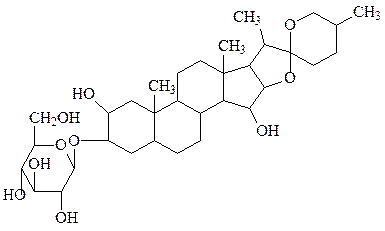
диготогенин- α-D-глюкозид (сердечный гликозид из наперстянки).
5. Этерификация – гидроксильные группы моносахаридов образуют эфиры с различными кислотами.
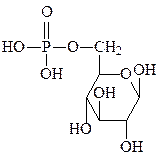
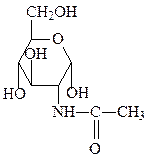
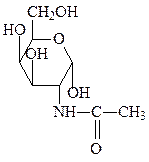
глюкозо-6-фосфат N-ацетилглюкозамин N-ацетилгалактозамин
Для анализа углеводов в растворе используют метод поляриметрии, который основан на способности оптически активных веществ вращать плоскость поляризованного света вправо или влево на угол α. Угол вращения 1М раствора D-глюкозы равен +52о.
Олигосахариды – это углеводы, состоящие из нескольких моносахаридов. В природе изредка встречаются трисахариды, имеющие в своем составе по три моносахарида, однако в основном распространены дисахариды.
Дисахариды – состоят из двух ковалентно связанных друг с другом моносахаридов. У восстанавливающих дисахаридов гликозидная связь между мономерами осуществляется за счет спиртового и полуацетального гидроксилов, поэтому такие дисахариды сохраняют один свободный полуацетальный гидроксил и все реакции, свойственные моносахаридам. У невосстанавливающих дисахаридов гликозидная связь образована за счет полуацетальных гидроксилов обоих моносахаридов, поэтому у них нет восстанавливающих свойств моносахаридов. Гликозидные связи легко гидролизуются кислотами, но устойчивы к действию щелочей. После того как моносахаридные единицы соединятся друг с другом, их называют остатками. Формулы основных дисахаридов приведены ниже.
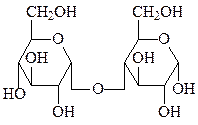
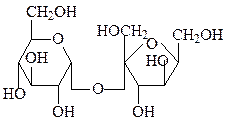
мальтоза сахароза
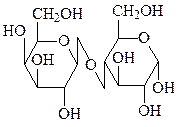
лактоза
Мальтоза образуется при расщеплении крахмала под действием амилаз и представляет собой α-D-глюкопиранозил-(1→4)-D-глюкопиранозид.
Лактоза является важнейшим углеводным компонентом молока млекопитающих и представляет собой β-D-галактопиранозил-(1→4)-D-глюкопиранозид.
Сахароза синтезируется только в растениях и служит в них растворимым резервным сахаром, который к тому же легко может транспортироваться. Устойчивость при транспорте и хранении обусловлена тем, что сахароза не является восстанавливающим сахаром, так как оба аномерных атома связаны друг с другом. Человека и животных сахароза привлекает своим сладким вкусом. По химическому строению это α-D-глюкопиранозил-(1↔2)-β-D-фруктопиранозид.
Полисахариды – содержат большое число моносахаридных остатков. Могут быть линейными и разветвленными.
Полисахариды можно разделить на два типа:
1) гомополисахариды (гомогликаны) – состоят из остатков одного и того же моносахарида – крахмал, гликоген;
2) гетерополисахариды (гетерогликаны) – содержат остатки двух и более моносахаридов – гиалуроновая кислота, муреин.
Важнейшие представители полисахаридов:
Крахмал – наиболее важный резервный полисахарид в клетках растений. Представляет собой смесь из двух полимеров глюкозы: α-амилозы и амилопектина. α-Амилоза состоит из длинных неразветвленных цепей, включающих 200-300 остатков D-глюкозы, соединенных друг с другом α(1→4) связями. Цепи амилопектина сильно разветвлены, в неразветвленных участках амилопектина 20-25 остатков глюкозы соединены друг с другом связями α(1→4), а в участках ветвления цепи – связями α(1→6). При этом формируется древовидная структура, в которой, как и в амилозе, имеется лишь одна свободная аномерная группа.
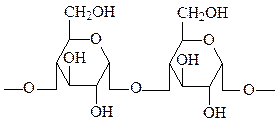
α-амилоза (15-20% крахмала)
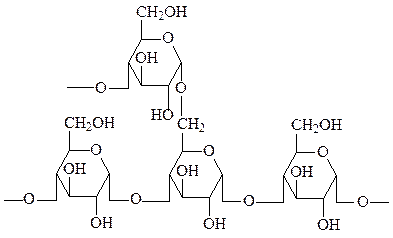
амилопектин в точке ветвления (80-85% крахмала)
В растениях крахмал содержится в хлоропластах листьев, плодах, семенах, клубнях. Особенно высоко содержания крахмала в зерновых культурах (до 75% от сухой массы) и клубнях картофеля (примерно 65%). Крахмал откладывается в специальных пластидах – амилопластах. Крахмальные гранулы практически не растворяются в холодной воде, однако они сильно набухают в воде при нагревании. При варке картофеля происходит экстракция амилозы горячей водой, в результате этого вода начинает опалесцировать и приобретает молочный оттенок. В вареном картофеле основную часть крахмала составляет оставшийся амилопектин. Некоторые виды крахмала, в частности крахмал «восковой» кукурузы, содержат один лишь амилопектин и не содержат амилозы. При неполном гидролизе крахмала образуются декстрины. Амилоза и амилопектин способны образовывать окрашенные комплексы с йодом синего и красного цвета соответственно.
Гликоген – основной резервный полисахарид в клетках животных. Аналогичен по строению амилопектину, только точки ветвления располагаются в среднем через каждые 8-10 остатков глюкозы, поэтому гликоген более разветвлен и компактен.
Целлюлоза – самое распространенное органическое соединение. Представляет собой линейный гомогликан, построенный из остатков глюкозы, связанных в положении β(1→4). Целлюлоза образует клеточные стенки растений и обладает высокой механической прочностью, устойчива к химическому и ферментативному гидролизу за счет образования водородных связей между параллельно лежащими молекулами, образующими фибриллы. Высшие животные не могут усваивать целлюлозу, однако у многих травоядных в желудочно-кишечном тракте содержатся симбиотические бактерии, способные расщеплять целлюлозу и переводить ее в форму, полезную для организма хозяина.
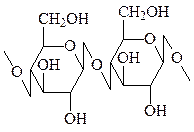
целлюлоза
Агароза – полисахарид из красных водорослей, представляет собой разветвленную цепь из галактозы и 3,6-ангидрогалактозы с β(1→4) типом связи, в точках ветвления β(1→3). Является водорастворимым полисахаридом, защищает водоросли от высыхания. Используется в микро-биологии как гелевая основа питательных сред (агар-агар).
Хитин – основной компонент наружного скелета насекомых и панциря ракообразных, также входит в состав клеточных стенок мицелия грибов. Представляет собой линейный полимер, образованный остатками N-ацетил-D-глюкозамина, которые связаны друг с другом β-связями. Хитиновый каркас у омаров и крабов усилен за счет включения карбоната кальция.
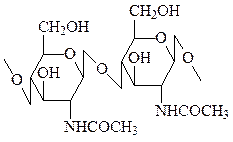
хитин
Декстран – компонент слизи некоторых бактерий, предохраняющий их от высыхания. Представляет собой полимер глюкозы, связанной преимущественно в положении α(1→6), а в точках ветвления в положении α(1→3). В воде декстран образует вязкие слизи или гели, из которых путем введения поперечных связей получают гидрофильные сорбенты для хроматографии – «сефадекс». Растворимый декстран находит применение в качестве заместителя плазмы при переливании крови.
ЛЕКЦИЯ 3
ЛИПИДЫ
Липиды – нерастворимые в воде маслянистые или жирные вещества, которые могут быть экстрагированы из клеток неполярными растворителями. Это гетерогенная группа соединений, непосредственно или опосредовано связанных с жирными кислотами.
Биологические функции липидов:
1) источник энергии, который может длительно запасаться;
2) участие в образовании клеточных мембран;
3) источник жирорастворимых витаминов, сигнальных молекул и незамени-мых жирных кислот;
4) теплоизоляция;
5) неполярные липиды служат электроизоляторами, обеспечивая быстрое распространение волн деполяризации вдоль миелинизированных нервных волокон;
6) участие в образовании липопротеидов.
Жирные кислоты – структурные компоненты большинства липидов. Это длинноцепочечные органические кислоты, содержащие от 4 до 24 углеродных атомов, они содержат одну карбоксильную группу и длинный неполярный углеводородный «хвост». В клетках не встречаются в свободном состоянии, а только в ковалентно связанной форме. В состав природных жиров входят обычно жирные кислоты с четным числом атомов углерода, поскольку они синтезируются из двууглеродных единиц, образующих неразветвленную цепь углеродных атомов. Многие жирные кислоты имеют одну или несколько двойных связей – ненасыщенные жирные кислоты.
Наиболее важные жирные кислоты (после формулы приведено число атомов углерода, название, температура плавления):
 12, лауриновая, 44,2оС
12, лауриновая, 44,2оС
 14, миристиновая, 53,9оС
14, миристиновая, 53,9оС
 16, пальмитиновая, 63,1оС
16, пальмитиновая, 63,1оС
 18, стеариновая, 69,6оС
18, стеариновая, 69,6оС
 18, олеиновая, 13,5оС
18, олеиновая, 13,5оС
 18, линолевая, -5оС
18, линолевая, -5оС
 18, линоленовая, -11оС
18, линоленовая, -11оС

20, арахидоновая, -49,5оС
Общие свойства жирных кислот;
- почти все содержат четное число атомов углерода,
- насыщенные кислоты у животных и у растений встречаются в два раза чаще, чем ненасыщенные,
- насыщенные жирные кислоты не имеют жесткой линейной структуры, они обладают большой гибкостью и могут принимать разнообразные конформации,
- в большинстве жирных кислот имеющаяся двойная связь расположена между 9-м и 10-м атомами углерода (Δ9),
- дополнительные двойные связи обычно расположены между Δ9-двойной связью и метильным концом цепи,
- две двойные связи в жирных кислотах не бывают сопряженными, между ними всегда находится метиленовая группа,
- двойные связи практически всех природных жирных кислот находятся в цис-конформации, что приводит к сильному изгибу алифатической цепи и более жесткому строению,
- при температуре тела насыщенные жирные кислоты находятся в твердом воскообразном состоянии, а ненасыщенные жирные кислоты представляют собой жидкости,
- натриевые и калиевые мыла жирных кислот способны эмульгировать нерастворимые в воде масла и жиры, кальциевые и магниевые мыла жирных кислот растворяются очень плохо и не эмульгируют жиров.
В мембранных липидах бактерий встречаются необычные жирные кислоты и спирты. Многие из бактериальных штаммов, содержащих эти липиды (термофилы, ацидофилы и галлофилы), адаптированы к экстре-мальным условиям.
 изоразветвленные
изоразветвленные
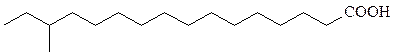 антеизоразветвленные
антеизоразветвленные
 циклопропансодержащие
циклопропансодержащие
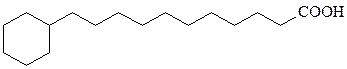 ω-циклогексилсодержащие
ω-циклогексилсодержащие
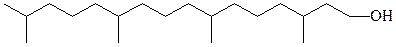 изопранильные
изопранильные
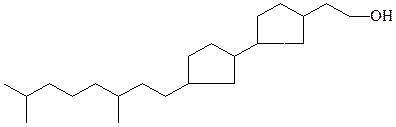 циклопентанфитанильные
циклопентанфитанильные
Состав бактериальных липидов отличается большим разнообразием и спектр жирных кислот разных видов приобрел значение таксономического критерия для идентификации организмов.
У животных важными производными арахидоновой кислоты являются гистогормоны простагландины, тромбоксаны и лейкотриены, объединенные в группу эйкозаноидов и обладающие чрезвычайно широкой биологической активностью.
 простагландин Н2
простагландин Н2
Классификация липидов:
1. Триацилглицериды (жиры) – это эфиры спирта глицерола и трех молекул жирных кислот. Они составляют основной компонент жировых депо растительных и животных клеток. В мембранах не содержатся. Простые триацилглицериды содержат остатки одинаковых жирных кислот во всех трех положениях (тристеарин, трипальмитин, триолеин). Смешанные содержат разные жирные кислоты. По удельному весу легче воды, хорошо растворимы в хлороформе, бензоле, эфире. Гидролизуются при кипячении с кислотами или основаниями, либо под действием липазы. В клетках в обычных условиях самоокисление ненасыщенных жиров полностью заторможено благодаря наличию витамина Е, различных ферментов и аскорбиновой кислоты. В специализированных клетках соединительной ткани животных адипоцитах огромное количество триацилглицеридов может запасаться в виде жировых капель, заполняющих почти весь объем клетки. В форме гликогена организм может запасти энергию не более чем на сутки. Триацилглицериды могут запасать энергию на месяцы, так как они могут храниться в очень больших количествах в практически чистом, негидратированном виде и в расчете на единицу веса в них запасается в два раза больше энергии, чем в углеводах. Кроме того, триацилглицериды под кожей образуют теплоизоляционный слой, защищающий организм от действия очень низких температур.
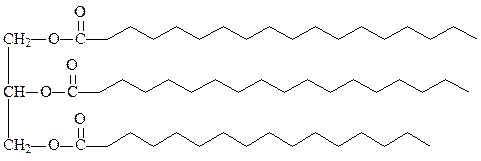 нейтральный жир
нейтральный жир
Для характеристики свойств жира используют следующие константы:
- кислотное число – количество мг КОН, необходимое для нейтрализации
свободных жирных кислот, содержащихся в 1 г жира;
- число омыления – количество мг КОН, необходимое для гидролиза
нейтральных липидов и нейтрализации всех жирных кислот,
содержащихся в 1 г жира;
- йодное число – количество граммов йода, связываемое с 100 г жира,
характеризует степень ненасыщенности данного жира.
2. Воска – это сложные эфиры, образуемые длинноцепочечными жирными кислотами и длинноцепочечными спиртами. У позвоночных животных секретируемые кожными железами воска выполняют функцию защитного покрытия, которое смазывает и смягчает кожу, а также предохраняет ее от воды. Восковым слоем покрыты волосы, шерсть, мех, перья животных, а также листья многих растений. Воска вырабатываются и ис-пользуются в очень больших количествах морскими организмами, особенно планктоном, у которого они служат основной формой накопления высококалорийного клеточного топлива.
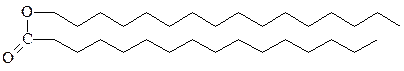
спермацет, получают из головного мозга кашалотов

пчелиный воск
3. Фосфоглицеролипиды – служат главными структурными компонентами мембран и никогда не запасаются в больших количествах. Обязательно содержат в своем составе многоатомный спирт глицерин, фосфорную кислоту и остатки жирных кислот.
Фосфоглицеролипиды по химическому строению можно еще разделить на несколько типов:
1) фосфолипиды – состоят из глицерина, двух остатков жирных кислот по 1-му и 2-му положению глицерина и остатка фосфорной кислоты, с которой связан остаток еще одного спирта (этаноламин, холин, серин, инозитол). Как правило, жирная кислота в 1-м положении насыщенная, а во 2-м – ненасыщенная.
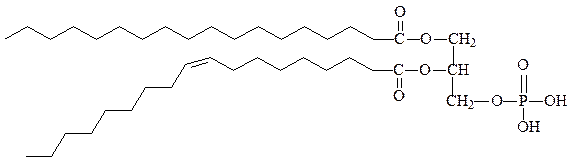
фосфатидная кислота – исходное вещество для синтеза других фосфолипидов, в тканях содержится в незначительных количествах
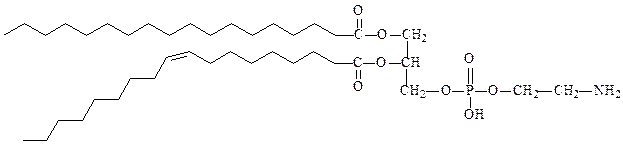 фосфатидилэтаноламин (кефалин)
фосфатидилэтаноламин (кефалин)
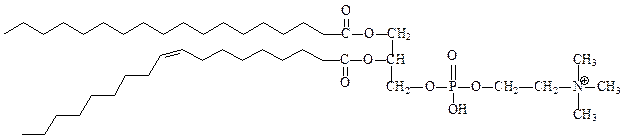 фосфатидилхолин (лецитин), его практически нет в бактериях
фосфатидилхолин (лецитин), его практически нет в бактериях
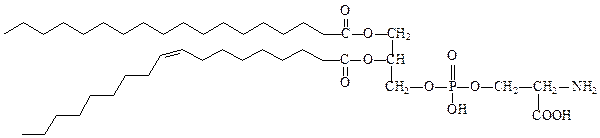 фосфатидилсерин
фосфатидилсерин
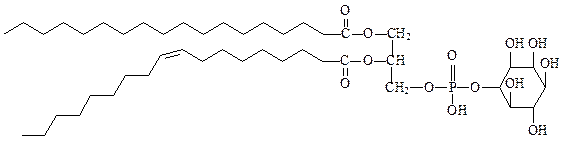
фосфатидилинозитол – предшественник двух важных вторичных мессенджеров (посредников) диацилглицерина и инозитол-1,4,5-трифосфата
2) плазмалогены – фосфоглицеролипиды, у которых одна из углеводо-родных цепей представляет собой простой виниловый эфир. Плазмалогены не встречаются в растениях. Этаноламиновые плазмалогены широко представлены в миелине и в саркоплазматическом ретикулуме сердца.
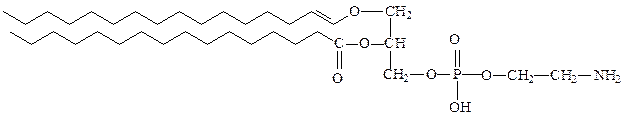 этаноламинплазмалоген
этаноламинплазмалоген
3) лизофосфолипиды – образуются из фосфолипидов при ферментатив-ном отщеплении одного из ацильных остатков. В змеином яде содержится фосфолипаза А2, которая образует лизофосфатиды, обладающие гемолитическим действием;
4) кардиолипины – фосфолипиды внутренних мембран бактерий и мито-хондрий, образуются при взаимодействии с глицерином двух остатков фосфатидной кислоты:
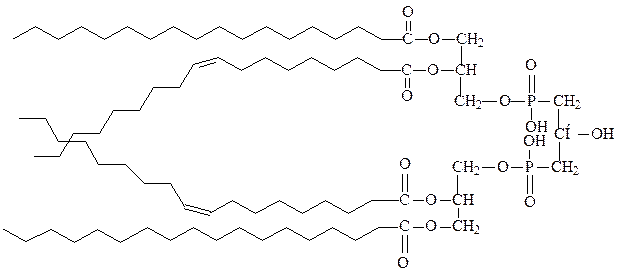 кардиолипин
кардиолипин
4. Фосфосфинголипиды – функции глицерина в них выполняет сфингозин – аминоспирт с длинной алифатической цепью. Не содержат глицерина. В большом количестве присутствуют в мембранах клеток нервной ткани и мозге. В мембранах растительных и бактериальных клеток фосфосфинголипиды встречаются редко. Производные сфингозина, ацилированного по аминогруппе остатками жирных кислот, называются церамидами. Важнейший представитель этой группы – сфингомиелин (церамид-1-фосфохолин). Он присутствует в большинстве мембран животных клеток, особенно много его в миелиновых оболочках нервных клеток определенного типа.
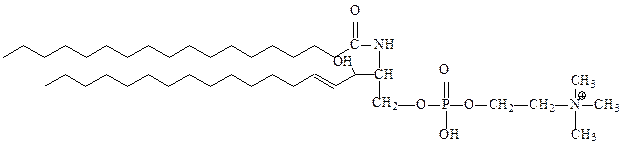 сфингомиелин
сфингомиелин
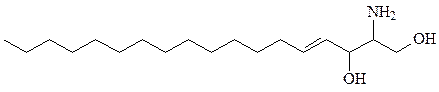 сфингозин
сфингозин
5. Гликоглицеролипиды –липиды, у которых в положении 3 глицерола находится углевод, присоединенный с помощью гликозидной связи, фосфатной группы не содержат. Гликоглицеролипиды широко представлены в мембранах хлоропластов, а также в сине-зеленых водорослях и бактериях. Моногалактозилдиацилглицерол – наиболее распространенный в природе полярный липид, поскольку на его долю приходится половина всех липидов тилакоидной мембраны хлоропластов:
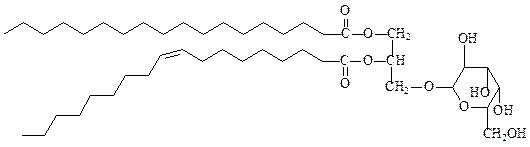
моногалактозилдиацилглицерол
6. Гликосфинголипиды– построены из сфингозина, остатка жирной кислоты и олигосахарида. Содержатся во всех тканях, главным образом в наружном липидном слое плазматических мембран. В них отсутствует фосфатная группа и они не несут электрического заряда. Гликосфинголипиды можно разделить еще на два типа:
1) цереброзиды – более простые представители этой группы. Галактоцереброзиды содержатся главным образом в мембранах клеток мозга, тогда как глюкоцереброзиды присутствуют в мембранах других клеток. Цереброзиды, содержащие два, три или четыре остатка сахаров, локализуются в основном в наружном слое клеточных мембран.

галактоцереброзид
2) ганглиозиды – наиболее сложные гликосфинголипиды. Их очень крупные полярные головы образованы несколькими остатками сахаров. Для них характерно наличие в крайнем положении одного или нескольких остатков N-ацетилнейраминовой (сиаловой) кислоты, несущей при рН 7 отрицательный заряд. В сером веществе головного мозга ганглиозиды составляют около 6% мембранных липидов. Ганглиозиды – важные компоненты расположенных на поверхности клеточных мембран специфических рецепторных участков. Так они находятся в тех специфических участках нервных окончаний, где происходит связывание молекул нейромедиатора в процессе химической передачи импульса от одной нервной клетки к другой.
7. Изопреноиды – производные изопрена (активная форма – 5-изопенте-нилдифосфат), выполняющие самые разнообразные функции.
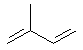 изопрен
изопрен 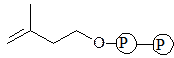 5-изопентенилдифосфат
5-изопентенилдифосфат
Способность синтезировать специфические изопреноиды свойственна лишь некоторым видам животных и растений.
1) каучук – синтезируют несколько видов растений, в первую очередь гевея бразильская:
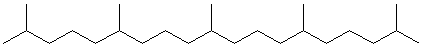 фрагмент каучука
фрагмент каучука
2) жирорастворимые витамины А, D, Е, К (из-за структурного и функ-ционального сродства со стероидными гормонами витамин D сейчас относят к гормонам):
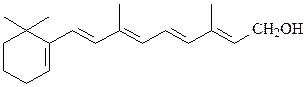 витамин А
витамин А
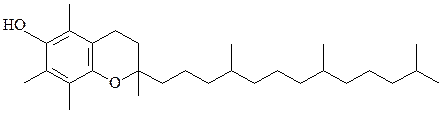 витамин Е
витамин Е
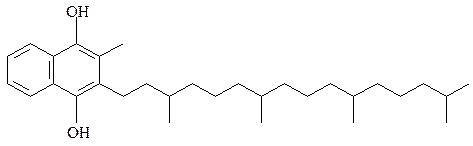 витамин К
витамин К
3) гормоны роста животных – ретиноевая кислота у позвоночных и неоте-нины у насекомых:
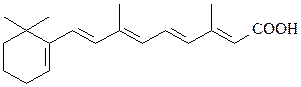 ретиноевая кислота
ретиноевая кислота
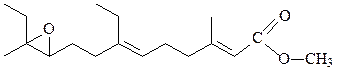 неотенин
неотенин
Ретиноевая кислота является гормональным производным витамина А, стимулирует рост и дифференциацию клеток, неотенины – гормоны насекомых, стимулируют рост личинок и тормозят линьку, являются антагонистами экдизону;
4) растительные гормоны – абсцизовая кислота, является стрессовым фитогормоном, запускающим системный иммунный ответ растений, проявляющийся в устойчивости к самым различным патогенам:
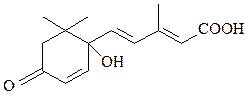 абсцизовая кислота
абсцизовая кислота
5) терпены – многочисленные душистые вещества и эфирные масла растений, обладающие бактерицидным и фунгицидным действием; соединения из двух изопреновых звеньев называются монотерпенами, из трех – сесквитерпенами, из шести – тритерпенами:
 камфора
камфора  тимол
тимол
6) стероиды – сложные жирорастворимые вещества, молекулы которых содержат в своей основе циклопентанпергидрофенантрен (по своей сути – тритерпен). Основной стерол в тканях животных – спирт холестерин (холестерол). Холестерин и его эфиры с длинноцепочечными жирными кислотами – важные компоненты липопротеинов плазмы, а также наружной клеточной мембраны. Из-за того, что четыре конденсированных кольца создают жесткую структуру, присутствие холестерина в мембра-нах регулирует текучесть мембран при экстремальных температурах. В растениях и микроорганизмах содержатся родственные соединения – эргостерин, стигмастерин и β-ситостерин.
 холестерин
холестерин
 эргостерин
эргостерин
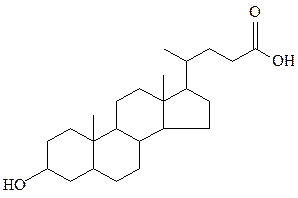 стигмастерин
стигмастерин
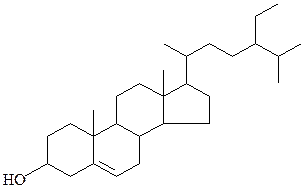 β-ситостерин
β-ситостерин
Из холестерина в организме образуются желчные кислоты. Они обеспечивают растворимость холестерина в желчи и способствуют перевариванию липидов в кишечнике.
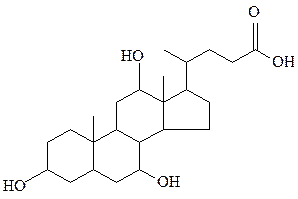 холевая кислота
холевая кислота
Из холестерина образуются также стероидные гормоны – липофильные сигнальные молекулы, регулирующие обмен веществ, рост и репродук-цию. В организме человека основными являются шесть стероидных гормонов:
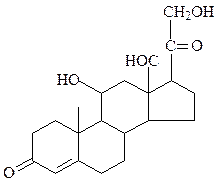
кортизол альдостерон
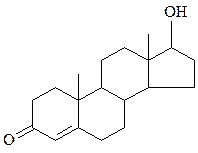

тестостерон эстрадиол
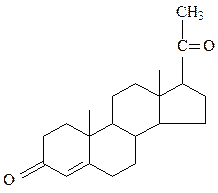
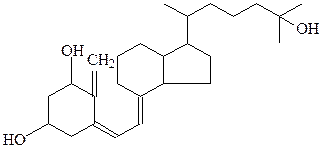
прогестерон кальцитриол
Кальцитриол – витамин D, обладающий гормональной активностью, он отличается от гормонов позвоночных, однако также построен на основе холестерина. Кольцо В раскрывается за счет светозависимой реакции.
Производным холестерина является гормон линьки насекомых, пауков и ракообразных – экдизон. Стероидные гормоны, выполняющие сигнальную функцию, встречаются также в растениях.
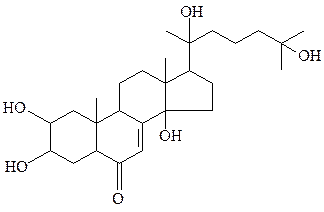 экдизон
экдизон
7) липидные якоря, удерживающие молекулы белков или других соединений на мембране:
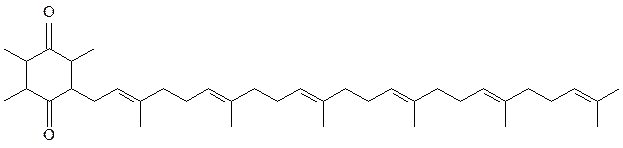 убихинон
убихинон
Как мы видим, липиды не являются полимерами в буквальном смысле этого слова, однако как в метаболическом, так и в структурном отношении они близки к присутствующей в бактериях полиоксимасляной кислоте – важному запасному веществу. Этот сильно восстановленный полимер состоит исключительно из звеньев D-β-оксимасляной кислоты, соединенных сложноэфирной связью. Каждая цепь содержит около 1500 остатков. Структура представляет собой компактную правую спираль, около 90 таких цепей уложено с образованием тонкого слоя в бактериальных клетках.
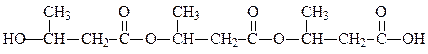
поли-D-β-оксимасляная кислота
ЛЕКЦИЯ 4
Дата добавления: 2015-08-08; просмотров: 2225;