Структура
Плотные контакты представлены в виде лент, состоящих из цепочек округлых зон (бляшек) размером 10 нм и с расстоянием между центрами соседних бляшек примерно 18 нм. Белки, составляющие многомолекулярный комплекс плотного контакта, можно разделить на три группы.
Интегральные белки, пересекающие апикальное межклеточное пространство и формирующие регулируемый барьер проницаемости. К ним относятся окклюдины, клаудины и молекула адгезионного контакта JAM (JunctionAdhesion Molecule).
Окклюдины (гены OCLN). Молекула белка проходит через плазмолемму 4 раза, образуя две внеклеточные петли и одну короткую цитоплазматическую петлю. Первая внеклеточная петля богата тирозином и глицином, соседняя петля содержит значительное количество заряженных аминокислотных остатков. COOH- и NH3-домены находятся в цитоплазматической части молекулы. Для COOH–конца характерно высокое содержание серина, треонина и тирозина, служащих мишенями для ряда протеин- и тирозинкиназ. Цитоплазматический домен окклюдина через белки zonula occludens ZO-1, ZO-2 и ZO-3 связан с актином кортикального цитоскелета (рис. 4-9). Окклюдины также участвуют в регуляции барьера параклеточной проницаемости.
Окклюдин в области плотного контакта находится в фосфорилированном состоянии. Дефосфорилированный окклюдин образует точечные локализации вдоль базолатеральной плазмалеммы. В стимулировании фосфорилирования окклюдина участвуют киназы — классическая протеинкиназа C (cPKC) и новая протеинкиназа C (nPKC), активированные диацилглицеролом.
Клаудины (гены CLDN). Клаудины составляют семейство из 24 белков 20–27 кД. Молекула клаудина, как и окклюдина, проходит через плазмолемму 4 раза. Аминокислотный состав двух внеклеточных петель существенно варьирует у разных клаудинов, давая в результате широкий размах изоэлектрических точек. COOH-конец молекулы клаудинов, за исключением клаудина-12, заканчивается участком, связывающимся с PDZ-доменом ZO-1, ZO-2 и ZO-3.
Дефекты CLDN3 могут быть причиной ряда сердечно-сосудистых и опорно-двигательных аномалий, наблюдаемых при синдроме Williams-Beuren (WBS) (синдром делеции смежного гена, затрагивающий гены хромосомы 7q11.23).
Молекула адгезионного контакта JAM(JAM-1–JAM-4) (F11R) относится к семейству иммуноглобулинов, Mr43 кД. JAM-1 проходит сквозь плазмолемму один раз и образует 2 внеклеточных V-петли; первая петля, имеющая NH2-конец, взаимодействует со своим гомологичным партнёром JAM-1 соседней клетки. Короткий внутриклеточный COOH-конец, содержащий PDZ-мотив, связывается с PDZ3-доменом ZO-1 и с PDZ9-доменом белка MUPP-1 (Multi-PDZ domain Protein 1).
Белки бляшки плотного контакта, многие из которых экспрессируют PDZ-домены. PDZ-Домен(PSd/SAP90,Discs Large, ZO-1) представляет собой последовательность из 80–90 аминокислот, формирующую своего рода гидрофобный «желобок», с которым связываются COOH–концы белков-мишеней. Белки бляшки плотного контакта являются связующими звеньями между интегральными белками и актиновыми филаментами, а также выступают в роли адаптеров при мобилизации молекул цитозоля, участвующих в межклеточных взаимодействиях.
PDZ-содержащие белки плотного контакта: белки ZO-1, ZO-2 и ZO-3, которые принадлежат семействуMAGUK (Membrane-Associated Guanylate Kinase); белки MAGI-1, MAGI-2, and MAGI-3 (Membrane-AssociatedGuanylyl kinase Inverted); MUPP-1; белки PAR-3 и PAR-6 (относящиеся к комплексу полярностиPAR3/aPKC/PAR6); белок PALS1 (Protein Associated Lin-7) и белок PATJ (PALS1-Associated Tight Junction). Оба белка относятся к семейству комплекса полярности Crumbs/PALS1/PATJ; белки комплекса полярностиScrib/mDgl/mLgl (Scrib-(Vartul)/mammalian discs large/mLgl).
Гетеротримерные G–белки образуют связи с ZO-1 и ZO-2. Как сигнальные молекулы участвуют в регуляции барьерной функции плотного контакта.
Сингулин (CGNL) — белок, связанный с JAM-1, ZO-1, ZO-2, ZO-3 плотного контакта. Формирует поперечные связи с белками плотного контакта, участвуя в образовании пучков нитей F–актина.
Разнородная группа цитоплазматических и ядерных белков, включая регуляторные белки, онкосупрессоры, факторы транскрипции, которые взаимодействуют, прямо или опосредованно, с белками бляшки плотного контакта для координации таких различных функций, как регуляция параклеточной проницаемости, клеточная пролиферация, полярность, онкосупрессия.
Симплекин (SYMPK) находится в составе бляшки плотного контакта совместно с ZO-1, а также присутствует в ядре. Участвует в процессинге пре-мРНК.
ZONAB (ZO-1-Associated Nucleic Acid Binding Protein) связан с ZO-1 в плотном контакте. Является фактором транскрипции Y-box; связывая CDK4, ZONAB регулирует клеточную пролиферацию, также может регулировать экспрессию ErbB2.
GEF-H1 (Guanine nucleotide Exchange Factor) — фактор, входящий в состав плотного контакта. Регулирует параклеточную проницаемость.
Rab3b, Rab13 привлекаются в состав адгезионных комплексов, формируемых при межклеточных контактах; могут участвовать в транспорте пузырьков к участкам межклеточной адгезии.
Sec6/Sec8 участвуют в транспорте пузырьков в области межклеточной адгезии.
Сборка
Сборка плотного контакта запускается гомофильным взаимодействием между E-кадгерином и нектином плазмолемм соседних клеток. Цитоплазматические домены нектина и E-кадгерина взаимодействуют между собой через комплекс афадин- катенин, при этом афадин связан с COOH–концом нектина, а катенин — с COOH–концом E-кадгерина. В ходе начальной стадии сборки JAM-1 соединяется с нектином (через белки, связанные сZO-1 и афадином); по окончании сборки JAM-1 и нектин разделяются. В ходе сборки контакта с JAM-1 могут связываться и другие цитоплазматические белки (сингулин, Ca2+/кальмодулин-зависимая сериновая протеинкиназаCASK). В регуляции сборки плотных контактов участвуют белки комплексов полярности PAR-3/aPKC/PAR-6,Crumbs3/PALS1/PATJ и Scrib/mDlg/mLgl. Белки комплексов полярности также играют центральную роль в асимметричном клеточном делении, в установлении передне-задней полярности зародыша в эмбриогенезе, организации апикально-базальной полярности эпителиальных клеток.
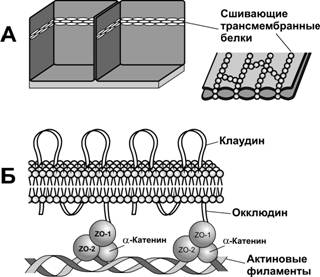
Рис. 4-9. Плотный контакт. (А)Сшивающие трансмембранные белки образуют непроницаемые для молекул цепочки локальных соединений между смежными плазматическими мембранами. (Б) В верхней части смежные клетки соединены при помощи клаудина, в нижней — окклюдина, длинный внутриклеточный домен которого соединён с белками ZO. Через белки ZO трансмембранные белки связаны с актиновыми филаментами цитоскелета. [17]
Дата добавления: 2015-07-07; просмотров: 1893;