Завершение антигеннезависимого лимфопоэза
а) Принципиальная особенность лимфопоэза
| Необхо-димо огромное множество BCR и ТCR | а) Завершающие стадии антигеннезависимого лимфопоэза проходят гораздо сложней, чем завершение любого направления миелопоэза. б) Дело в том, что зрелые лимфоциты обладают иммуноспецифичностью – содержат на поверхности белковые рецепторы (см. тему 8, пункт 7). - У В-лимфоцитов это BCR (состоят из двух разных полипептидных цепей), способные узнать тот или иной антиген. - У Т-лимфоцитов – TCR (по одной полипептидной цепи), настроенные на определённую антигенную детерминанту (АД, т.е. пептид из 8–15 аминокислотных остатков) – при условии, что АД связана со специальным белком и выставлена на поверхности клеток. |
| Но генов на все BCR, ТCR не напасёшься | Так вот, сложность обусловлена тем, что в геноме и ГСК (гемопоэтических стволовых клеток), и всех прочих клеток организма, нет и не может быть сразу всего того огромного множества генов, которые кодировали бы - BCR ко всем возможным антигенам и - TCR ко всем возможным АД. |
| И всё-таки они образуются! | а) Тем не менее, при созревании лимфоцитов подобные множества формируются – но только не в одной клетке, а в огромном количестве клеточных клонов, где - Т-клетки одного клона содержат ген (гены) только для одного ТCR - и В-клетки одного клона – гены только для одного BCR. б) Образование этих клонов и генов для TCR и BCR происходит в ходе т.н. реаранжировкигенов и последующей селекцииклеток. |
б) Реаранжировка генов в лимфопоэзе
| Исход-ный генети-ческий материал | а) В хромосомах ГСК (и любой другой клетки) содержатся - не длинные (полные) гены целых цепей BCR и TCR всевозможной иммуноспецифичности, - а относительно короткие гены нескольких полипептидных участков, из которых состоит цепь BCR и цепь TCR, б) Причём, для каждого такого участка имеется до нескольких сотен вариантов (аллелей) его короткого гена, и все они вместе присутствуют в геноме. в) Если условно считать, что - для формирования двух полных генов BCR надо четыре коротких, - и у каждого короткого гена имеется по 100 аллелей, то всего в геноме за структуру BCR отвечает менее 500 коротких генов. |
| Собственно реаранжировка | а) В ходе реаранжировки генов специальные ферменты (рекомбиназы) - «выхватывают» из каждого аллельного множества по одному произвольно выбранному короткому гену и - «сшивают» их в определённом порядке в два полных гена BCR. б) Тогда общее число вариантов образующихся затем BCR равно 100 × 100 × 100 × 100 ≈ 108 . в) Т.к. в каждой клетке реализуется лишь один вариант, то столько же клонов В-лимфоцитов может появиться в результате реаранжировки генов. |
в) Селекция клеток
| Уцелев-шие клетки необходимо проверить! | а) Конечно, во многих клетках перестройка генома приводит к неисправимым повреждениям хромосом, что инициирует в клетках программу самоуничтожения (апоптоза). б) Сохранившиеся клетки размножаются, образуя клоны с разной иммунной специфичностью. в) Но до выхода клеток в сосудистое русло должна происходить их селекция. Наиболее изучена селекция Т-лимфоцитов в корковом веществе долек тимуса. |
| Положи-тельная селекция | а) Первый тур – положительная селекция: отбираются клетки, способныевзаимодействовать своими рецепторами (TCR) и корецепторами с антигенпредставляющими клетками. б) Клетки, способные к эффективному взаимодействию, сохраняются. В других клетках инициируется апоптоз. |
| Отрица-тельная селекция | а) Второй тур – отрицательная селекция: теперь клетки проверяются на способность взаимодействовать с АД собственных белков организма. б) Эти АД им представляют специализированные макрофаги (интердигитирующие и дендритные клетки). в) Теперь апоптоз инициируется в клетках, способных к указанному взаимодействию. г) И сохраняются клетки, дающие отрицательный результат. Им-то и предоставляется возможность выйти в кровеносное русло и стать циркулирующим лимфоцитом. |
6. Красный костный мозг (ККМ)а) Состав
| Компо-ненты | Кроветворные органы (в т.ч. и ККМ) содержат 4 компонента: 1) стромальный, 2) гемальный (гемопоэтические клетки), 3) клетки макрофагического ряда и 4) специфические сосуды |
| 1) Строма | а) Строма ККМ – ретикулярная ткань, включающая ретикулярные клетки и ретикулярные волокна (из коллагена III типа). В ячейках стромы и лежат кроветворные клетки. б) Кроме того, к стромальному компоненту ККМ относят клетки ещё трёх видов: остеогенные (в составе эндоста), адвентициальные (на поверхности сосудов) и жировые. |
| 2) Гемальный компонент | а) Очевидно, в ККМ находятся почти все те гемопоэтические клетки, которые были показаны на рис. 20.1. б) При этом они расположены островками – эритропоэтическими, гранулоцитопоэтическими и т.д., – в которых преобладают клетки класса V. |
| 3) Мак-рофаги | В ККМ – три вида макрофагов: типичные, остеокласты (в составе эндоста) и т.н. клетки-кормилки. Последние поглощают железо из крови и передают его эритробластам. |
| 4) Сосуды | а) Сосуды подходят к ККМ из костного вещества. б) Среди капилляров ККМ одни (трофические) остаются узкими. Другие переходят в широкие синусные капилляры – место выхода клеток из ККМ. |
б) Препарат
| Локали-зация | а) Препарат, представленный на снимке, приготовлен из эмбриона. б) Поэтому ККМ находится междуформирующимисякостнымибалками (5). |
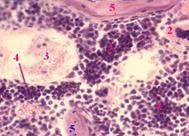 Рис.20.2. Срез красного костного мозга
Окраска гематоксилином и эозином
Рис.20.2. Срез красного костного мозга
Окраска гематоксилином и эозином
|
| Гемаль-ный компо-нент | а) ККМ выглядит как скопление множества клеток с тёмно-фиолетовыми ядрами. б) Это в основном гемопоэтические клетки (1) разных стадий развития. в) Различить среди них отдельные виды клеток практически невозможно. | |
| г) Исключение составляют мегакариоциты (2) – очень крупные многоядерные клетки с оксифильной (розовой) цитоплазмой. | ||
| Прочие компо-ненты | а) Видны также синусоидные сосуды (3), выстланные плоскими эндотелиоцитами (4). б) Клетки стромы (главным образом, ретикулярные клетки) имеют неправильную форму, светло-розовую цитоплазму и бледноокрашенное ядро. |
7. Тимуса) Общие сведения
| Дольки и их части | а) Тимус покрыт капсулой (1) и отходящими от неё перегородками (2) разбит на дольки (3). б) В каждой дольке различают - на периферии – корковое вещество (4), плотно заселённое лимфоцитами и потому на препарате более тёмное, - и в центре – мозговое вещество (5), более светлое. |
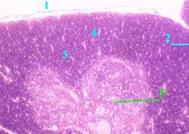 Рис. 20.3
Тимус ребёнка
Окраска гем. и эозином
Рис. 20.3
Тимус ребёнка
Окраска гем. и эозином
|
| Строма | Одна из особенностей тимуса – в том, что его строма имеет эпителиальную природу: | |
| представлена не ретикулярными клетками и одноимёнными волокнами, а эпителиоретикулярными клетками (без волокон). |
б) Характеристика коркового и мозгового вещества
| КОРКОВОЕ ВЕЩЕСТВО | МОЗГОВОЕ В-ВО | |
| Лимфоидные клетки | а) В подкапсулярной области – Т-лимфобласты (клетки класса IV). б) Глубже – созревающие Т-клетки, проходящие реаранжировку генов и два тура селекции. | а) В мозговом веществе – рециркулирующие Т-лимфоциты, попадающие в кровь и обратно через т.н. венулы с высоким эндотелием. б) Клетки стромы могут образовывать слоистые тельца Гассаля – округлые наслоения ороговевших эпителиоцитов. |
| Клетки стромы | Функциональные виды стромы: 1) опорные клетки, в т.ч. те, что отростками формируют гематотимусныйбарьер, 2) клетки-«няньки»; в их инвагинациях развиваются Т-лимфоциты; 3) секреторные клетки. | |
| Макрофаги | Обычные макрофаги, дендритные и интердигитирующие клетки. |
Тема 21. КРОВЕТВОРЕНИЕ: ПЕРИФЕРИЧЕСКИЕ ОРГАНЫ КРОВЕТВОРЕНИЯ И ИММУНОГЕНЕЗА
| Суть и механизм иммунных реакций в кратком виде обсуждались в теме 8 (пункт 7). Здесь мы расширим приведённую там информацию и, главное, «привяжем» происходящие события к конкретным участкам органов иммуногенеза. |
1. Общие сведенияа) Функция органов иммуногенеза
| Трой-ной барьер | Эти органы образуют систему барьеров для антигенов: а) первый барьер – лимфоидная система слизистых оболочек, б) второй барьер – множество лимфоузлов по ходу лимфатических сосудов: задерживают антигены, проникшие через кожу и слизистые оболочки в лимфу; в) третий барьер – селезёнка: задерживает антигены, оказавшиеся в крови. |
| Резуль-тат иммунной реакции | а) Причём, задержание антигена инициирует в органах иммуногенеза иммуннуюреакцию. б) Результатом её является формирование эффективной системы поиска и уничтожения данного антигена не только в отдельном локусе, но и по всему организму: - в случае клеточной иммунной реакции – в виде множества активных Т-киллеров, настроенных на уничтожение вполне определённого «врага», - а в случае гуморальной иммунной реакции – в виде не менее эффективных антител, способных найти и нейтрализовать соответствующий антиген почти в любой точке организма. |
б) Объекты узнавания
| Условие реакции на антиген | а) Ключевой принцип иммунных реакций – принцип двойного узнавания: агент может быть признан чужеродным и соответствующим образом атакован, только если какая-либо его антигенная детерминанта (АД) или полный антиген опознаны сразу двумя «свидетелями»: В-клеткой и Т-хелпером либо Т-киллером и Т-хелпером. б) В связи с этим, уточним, какие объекты узнаются этими клетками. | ||
| Что узнают Т-клетки – киллеры и хелперы | а) В-лимфоциты узнают АД в нативных целых макромолекулах – свободных или на поверхности клетки. б) А для Т-лимфоцитов требуется выполнение 3-х условий: АД должна быть 1) выделена из состава белка, 2) связана со специальным белком-демонстратором и 3) выведена на поверхность клетки. в) Но для Т-хелперов и Т-киллеров и здесь есть большие различия. | ||
| Белки-демон-страторы Локализация этих белков Представляемые ими АД | Объекты, узнаваемыеТ-киллером Антигены ГКГ (главного комплекса гистосовместимости) I-го класса На поверхности почти всех клеток организма (в т.ч. и АПК) АД эндогенных (синтезированных в данной клетке) белков | Объекты, узнаваемыеТ-хелпером Антигены ГКГ II-го класса На поверхности только АПК (антигенпредставляющих клеток) АД внешних антигенов, поглощённых и переработанных | |
в) Как образуются объекты, узнаваемые Т-клетками
| Эндогенная АД – антиген ГКГ-I Многие эндогенные белки распадаются впротеосомах до пептидов по 8–11 остатков аминокислот (АК). Пептиды связываются с белками ГКГ-I и выводятся на поверхность клетки. | Экзогенная АД – антиген ГКГ-II Белки, поглощённые путём фагоцитоза антигенных объектов, распадаются в лизосомах до пептидов по 15–20 остатков АК. Пептиды связываются с белками ГКГ-II и тоже выводятся на поверхность клетки. |
Дата добавления: 2015-08-04; просмотров: 1130;