Механізм злиття клітин
Для індукції злиття клітин використовуються речовини різної природи. Іони Са2+, поліетиленгліколь, лізолецитин, моноолеат гліцерину, вірус Сендай.
Лізолецитин – поверхнево-активна речовина ліпідної природи, продукт деградації лецитину шляхом обробки останнього фосфоліпазою А. Лізолецитин пошкоджує мембрани й токсичний для живих систем. Цитотоксичний ефект цієї речовини можна зменшити знижуючи його концентрацію або додаючи під час обробки альбумін.
Моноолеат гліцерину також сполука ліпідної природи, але її ушкоджуюча дія менш виражена, а частота злиття клітин при застосуванні цієї речовини зростає в 4 – 7 разів у порівнянні зі спонтанним процесом. До інших аглютинуючих агентів, здатність яких викликати злиття клітин, була досліджена спеціально, відносяться лектини рослин й антитіла.
Перевага вірусу Сендай як агрегуючого агенту, полягає в повній відсутності цитотоксичного ефекту. Вірус перед уживанням інактивують, опромінюючи ультрафіолетовою лампою протягом 5 хвилин, при цьому він втрачає здатність до розмноження, але зберігає здатність зливати клітини. Вірус Сендей має два недоліки:
– необхідність нарощувати, титрувати, концентрувати й інактивувати вірус;
– клітини рослин і грибів не мають рецепторів до цього вірусу, тому він не застосовується для їхньої гібридизації.
Перший етап злиття (рис. 6) – зближення мембран сусідніх клітин і встановлення між ними тісного контакту. Мембрани повинні бути наближені один до одного на відстані у декілька ангстрем так, щоб між ними були можливі взаємодії, подібні до гідрофобних зв’язків. Викликають подібне зближення агенти, які індукують аглютинацію клітин. Міксовіруси, наприклад, вірус Сендай, поряд з іншими вірусами, які не викликають злиття, насамперед викликають аглютинацію клітин, тобто досить тісне їхнє зближення, необхідне для успішного наступного злиття.
Поліетиленгліколь (ПЕГ) також викликає агрегацію клітин, хоча механізм дії його невідомий. Можливо, завдяки тому, що у водному розчині ПЕГ несе невеликий негативний заряд, молекули цього розміру досить великі, щоб між клітинами виникали електростатичні зв’язки. Підтвердженням цієї гіпотези є посилення аглютинації клітин, що викликається ПЕГ: двовалентні іони, очевидно, утворюють містки між ПЕГ і негативно зарядженими вуглеводами, що перебувають на клітинній поверхні. Відповідно до іншої гіпотези, протопласти зливаються в результаті дегідратації. Поглинання води індукує утворення пор на поверхні мембрани й відбувається перетікання внутрішньоклітинного матеріалу. Після злиття ділянки з порами зберігаються якийсь час. Існує два припущення, що пояснюють виникнення пор :
1. При високій концентрації ПЕГ (20 – 30%) вся вільна вода поглинається ним, викликаючи розриви в мембрані;
2. ПЕГ зменшує полярність води, що викликає перерозподіл полярних і гідрофобних компонентів мембрани, що стабілізують ліпідні шари.
З особливим успіхом для цих цілей використовується ПЕГ з молекулярною масою від 1500 до 7500.
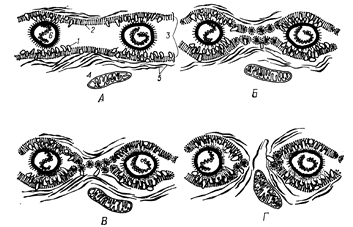
Рис. 6. Етапи злиття клітин (за Н. Рінгертцом, Р. Севіджу, 1979): А – 1-й етап, зближення цитоплазматичних мембран: 1 – глікопротеїди, 2 – ліпіди, 3 – плазматичні мембрани, 4 – мітохондрії, 5 – мікрофіламенти, 6 – частки вірусу Сендай; Б – 2-й етап, вихід глікопротеїдів й оголення ліпідних шарів мембрани; В – 3-й етап, утворення міцел; Г – початок 4-го етапу, злиття мембран, утворення цитоплазматичних містків.
Лектини й антитіла – двох- або полівалентні сполуки, і їхня аглютинуюча функція пов’язана зі здатністю однієї молекули якого-небудь із цих сполук взаємодіяти з рецепторами, що перебувають на поверхні двох клітин, що й приводить до утворення зв’язку між клітинами. Достатня кількість таких молекулярних зв’язків може втримувати клітини разом, перешкоджаючи їхньому розходженню або в результаті броунівського руху, або в результаті електростатичного відштовхування, або в результаті активної міграції клітин.
На другому етапі глікопротеїди, розташовані на поверхні, починають вивільнятися з ділянок мембрани, що лежать між віріонами, і притягуються до місць прикріплення вірусних часток. Вуглеводні компоненти – найменш вивчена частина клітинної поверхні. У мембранах вони зустрічаються у вигляді нейтральних цукрів, а також ковалентно зв’язуються з ліпідами або білками. Саме в такому вигляді вони беруть активну участь у багатьох біологічних процесах. Встановлено, що глікопротеїди визначають антигенну специфічність клітин, несуть негативний заряд, характерний для клітин при фізіологічно нейтральних рН, беруть участь у розпізнаванні й адгезії клітин, визначають рецепторні ділянки для вірусів, бактерій, аглютинуючих агентів, беруть участь у процесі регуляції проникності мембран для іонів. Ці властивості й визначають їхню участь у злитті клітин.
Вуглеводи перешкоджають злиттю клітин, тому що розділяють ліпідні шари мембран, не даючи їм стикатися, тому що на поверхні нормальної клітини рецепторні ділянки розташовані рідко або поодинці. У результаті трансформації вірусом відбувається їхнє об’єднання в групи. Глікопротеїди мігрують у мембрані до місця адсорбції вірусу, залишаючи вільними сусідні ділянки, де й відбувається злиття клітин. У деяких випадках у видаленні вуглеводних груп, які перешкоджають злиттю клітин, беруть участь лізосомні ферменти. При цьому відбувається з’єднання лізосом або пухирців АГ із плазматичною мембраною, і локальне вивільнення глікозидаз. Така лізосомна активність може бути запрограмована (у гаметах, міобластах, макрофагах, тобто клітинах, що зливаються в природних умовах) або викликана експериментально при індукованому злитті клітин.
Третій етап – міцелізація оголених ліпідів двох протилежних мембран. Міцели – ліпідні краплі, де молекули ліпідів гідрофільними голівками обернені до води, а гідрофобні “хвости” жирних кислот заховані всередину. Цей процес підсилюється при високих значеннях рН і високій концентрації іонів кальцію. В обох клітинах починається ендоцитоз вірусних часток.
Четвертий етап – злиття мембран. Завдяки місткам, утвореним Са2+, розмір міцел зменшується. Під дією АТФ і Са2+, активізуються мікрофіламенти. Виниклий цитоплазматичний місток стабілізується й розширюється завдяки функціонуванню мікрофіламентів. На останніх етапах злиття клітин важливу роль відіграє наявність АТФ. АТФ утворюється завдяки діяльності мітохондрій. Полікаріони, що утворюються при злитті двох або трьох клітин, являють собою не безформні роздуті або дволопатеві кулі цитоплазми, а приймають форму, характерну для однієї з батьківських клітин або проміжну між ними. Форму клітини визначають мікротрубочки й мікрофіламенти. У процесі злиття клітин мікрофіламенти актиноміозинового типу, розташовані під цитоплазматичною мембраною, беруть безпосередню участь, тому що забезпечують з’єднання цитоплазм клітин, що злилися, і стабілізацію новоутвореної системи. Однак, формування цитоскелету – енергозалежний процес, який вимагає великої кількості АТФ. Напевно, цим обумовлена присутність мітохондрій у місцях злиття клітинних мембран.
Дата добавления: 2015-07-14; просмотров: 1179;