Утворення енергії внаслідок окиснення глікогену
| Етап процесу | АТФ, утворюваний з 1 моля глікогену печінки | |
| безпосередньо | окиснювальним фосфорилюванням* | |
| Гліколіз (перетворення глюкози на піровиноградну кислоту) | ||
| Перетворення піровиноградної кислоти на ацетил-КоА | ||
| Цикл Кребса | 
| 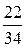
|
| Усього | ||
| * Відноситься до АТФ, утвореного при переносі Н+ електронів у ланцюжок переносу електронів. |
Окиснення жирів. Як уже відмічалося, жири також роблять свій внесок в енергетичні потреби м'язів. Запаси глікогену у м'язах та печінці можуть забезпечити усього 1200-2000 ккал енергії, у той час як жири, що містяться всередині м'язових волокон та в жирових клітинах, — близько 70 000-75 000 ккал.
Хоча жирами називають багато хімічних сполук, такі, як тригліцериди, фосфоліпіди та холестерин, тільки тригліцериди використовуються як основні джерела енергії. Тригліцериди знаходяться у жирових клітинах та волокнах скелетних м'язів. Щоб використати тригліцериди для утворення енергії, необхідно розщепити їх на основні складові: одну молекулу гліцерину та три молекули вільних жирних кислот. Цей процес називається ліполізом і здійснюється ферментами — ліпазами. Оскільки вільні жирні кислоти — це основне джерело енергії, ми приділимо їм головну увагу.
Вивільнившись з тригліцериду (жиру), вільні жирні кислоти можуть попасти у кров, котра транспортує їх по усьому організму, і у результаті дифузії проникнути у м'язові волокна. Інтенсивність надходження вільних жирних кислот у м'язові волокна залежить від градієнта концентрації. Підвищення концентрації вільних жирних кислот у крові виштовхує їх у м'язові волокна.
Процес β-окиснення. Незважаючи на значні структурні відмінності між різними вільними жирними кислотами, їх метаболізм майже однаковий, як видно на рис. 4.8. До того, як вони попадуть у м'язові волокна, вільні жирні кислоти активуються енергією АТФ за допомогою ферментів. Таким чином їх готують до катаболізму (розщеплення) у мітохондріях. Цей ферментний катаболізм жирів мітохондріями називається β-окисненням.
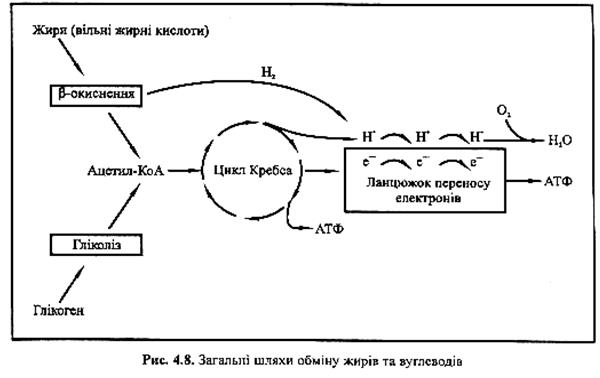
У цьому процесі вуглецевий ланцюжок вільної жирної кислоти ділиться на двовуглецеві рештки оцтової кислоти. Наприклад, якщо первісне вільна жирна кислота мала 16-вуглецевий ланцюжок, то при β-окисненні утворюється 8 молекул оцтової кислоти. Вся оцтова кислота потім перетворюється на ацетил-КоА.
Цикл Кребса та ланцюжок переносу електронів. З цього моменту обмін жирів здійснюється за тим же принципом, що й метаболізм вуглеводів. Ацетил-КоА, що утворився внаслідок β-окиснення, вступає у цикл Кребса. У цьому циклі утворюється водень, котрий транспортується у ланцюжок переносу електронів разом з воднем, утвореним під час β-окиснення, де піддається окиснювальному фосфорилюванню. Як і при обміні глюкози, проміжними продуктами окиснення вільних жирних кислот є АТФ, Н2О, СО2. Однак для повного спалювання молекули вільних жирних кислот потребується більше кисню.
Хоча жири забезпечують більше кілокалорій енергії на грам, ніж вуглеводи, для їх окиснення лотребується більше кисню, ніж для окиснення вуглеводів. Жири утворюють 5,6 молекули АТФ відносно молекули О2 вуглеводи — 6,3 молекули АТФ відносно молекули О2. Доставка кисню обмежена кисневотранспортною системою, тому кращим джерелом енергії під час виконання фізичної вправи високої інтенсивності є вуглеводи.
Переважний вміст у вільних жирних кислотах більшої кількості вуглецю, ніж у глюкозі, полягає в утворенні більшої кількості ацетил-КоА при метаболізмі даної кількості жиру, отже, у цикл Кребса надходить більше ацетил-КоА, а у ланцюжок переносу електронів — більше електронів. Саме тому при метаболізмі жирів утворюється набагато більше енергії, ніж при метаболізмі вуглеводів.
Таблиця 4.3
Утворення енергії внаслідок окиснення пальмітинової кислоти (С16Н32О2)
| Етап процесу | АТФ, утворюваний з 1 молекули пальмітинової кислоти | |
| безпосередньо | окиснювальним фосфорилюванням | |
| Активація жирної кислоти | - | |
| β-окиснення | - | |
| Цикл Кребса | 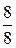
| 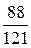
|
| Усього |
Візьмемо, наприклад, пальмітинову кислоту — досить поширену 16-вуглецеву вільну жирну кислоту. В результаті реакцій окиснення, циклу Кребса та ланцюжка переносу електронів з однієї молекули пальмітинової кислоти утворюється 129 молекул АТФ (табл. 4.3), у той часяк з молекули глюкози та глікогену відповідно 38 та 39 молекул. Незважаючи на такий високий показник, тільки близько 40 % енергії, що вивільнюється внаслідок метаболізму молекул або глюкози, або вільних жирних кислот, витрачається на утворення АТФ. Решта 60 % виділяються у вигляді тепла.
МЕТАБОЛІЗМ БІЛКІВ
Як уже відмічалося, вуглеводи та жири є переважними джерелами енергії нашого організму. Однак використовуються і білки або, скоріше, амінокислоти, з котрих вони складаються. Деякі амінокислоти можуть перетворюватися на глюкозу (за допомогою глюконеогенезу). Інші можуть перетворюватися на різні проміжні продукти окиснювального метаболізму (такі, як піруват або ацетил-КоА), щоб взяти участь в окислювальному процесі.
Кількість енергії, утворюваної білками, досить важко визначити, на відміну від енергії, утворюваної вуглеводами або жирами, оскільки білки також містять азот. При катаболізмі амінокислот певна кількість азоту використовується для утворення нових амінокислот, решта азоту перетворюється на сечовину і виділяється головним чином з сечею. Цей процес потребує використання АТФ і, отже, призводить до витрат якоїсь кількості енергії.
При розщепленні білків шляхом спалювання у лабораторних умовах утворюється 5,65 ккал∙г-1 енергії. При метаболізмі білка в організмі, внаслідок витрат енергії на процес перетворення азоту на сечовину, вивільнюється усього 5,20 ккал∙г-1 енергії, тобто на 8 % менше.
Щоб точно визначити інтенсивність обміну білків, необхідно встановити, яка кількість азоту виводиться з організму. Для цього необхідно збирати сечу протягом 12-24 год. Оскільки здоровий організм використовує невелику кількість білків у стані спокою і при виконанні фізичного навантаження (як правило, набагато менше 5-10 % усіх витрат енергії), при оцінці витрат енергії метаболізм білків просто не беруть до уваги.
ОКИСНЮВАЛЬНІ ЗДАТНОСТІ М'ЯЗІВ
Ми з'ясували, що процеси окиснювального метаболізму забезпечують максимальне утворення енергії. Було б ідеальним, якби ці процеси завжди здійснювалися з максимальною ефективністю. Однак,як і усі фізіологічні системи, вони мають свої обмеження. Окислювальна здатність Qо2 м'яза — це показник її максимальної здатності використовувати кисень. Далі ми розглянемо чинники, що обмежують цю здатність м'язів.
Активність ферментів. Здатність м'язових волокон окиснювати вуглеводи та жири досить важко визначити. У багатьох дослідженнях спостерігали тісний взаємозв'язок між здатністю м'яза виконувати аеробну вправу протягом тривалого часу та активністю її окиснювальних ферментів. Оскільки для окиснювання потребується багато ферментів, то їх активність у м'язових волокнах є достатньо надійним показником окиснювального потенціалу.
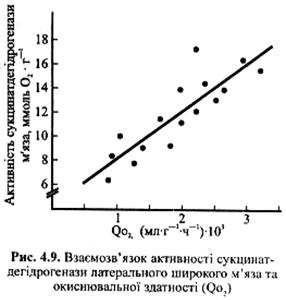
Немає рації вимірювати усі ферменти, тому для ілюстрації аеробної здатності волокон вибирають деякі з них. Найчастіше використовують такі ферменти, як сукцинатдегідрогеназу та цитратсинтазу, ферменти мітохондрій, що беруть участь у циклі Кребса. На рис. 4.9 проілюстровано взаємозв'язок між активністю сукцинатдегідрогенази у латеральному широкому м'язі та її окиснювальною здатністю. Окиснювальна активність ферментів у м'язах спортсменів, які займаються видами спорту, що потребують прояву витривалості, у два-чотири рази вища, ніж у м'язах нетренованих чоловіків та жінок.
 Склад м'язових волокон та тренувальні навантаження, спрямовані на розвиток витривалості. Склад волокон м'яза частково визначає його окиснювальну здатність. Як уже відмічалося у розділі 2, ПС волокна мають більшусхильність до аеробної діяльності, ніж ШС, оскільки містять більше мітохондрій та окиснювальних ферментів. ШС волокна більш придатні для гліколітичного продукування енергії. Отже,чим більше у м'язах ПС волокон,тим вища їх окиснювальна здатність.Наприклад, у найсильніших бігунівна довгі дистанції значно більше ПСволокон, мітохондрій і вища активність окиснювальних ферментів, ніжу нетренованих людей.
Склад м'язових волокон та тренувальні навантаження, спрямовані на розвиток витривалості. Склад волокон м'яза частково визначає його окиснювальну здатність. Як уже відмічалося у розділі 2, ПС волокна мають більшусхильність до аеробної діяльності, ніж ШС, оскільки містять більше мітохондрій та окиснювальних ферментів. ШС волокна більш придатні для гліколітичного продукування енергії. Отже,чим більше у м'язах ПС волокон,тим вища їх окиснювальна здатність.Наприклад, у найсильніших бігунівна довгі дистанції значно більше ПСволокон, мітохондрій і вища активність окиснювальних ферментів, ніжу нетренованих людей.
Тренувальні навантаження, спрямовані на розвиток витривалості, збільшують окиснювальні здатності усіх волокон і особливо ШС, ставлячи високі вимоги до окиснювального фосфорилювання, вони стимулюють м'язові волокна до утворення великої кількості мітохондрій, що містять велике число окиснювальних ферментів. Збільшуючи кількість ферментів у волокнах для β-окиснення, такі навантаження також допомагають м'язам більшою мірою розраховувати на жири як джерело продукування АТФ.
Таким чином, тренувальні навантаження на розвиток витривалості дають змогу підвищити аеробні здатності м'язів навіть у людей з високим вмістом ШС волокон. Разом з тим відомо, що ШС волокно у результаті тренування на розвиток витривалості не зможе такою ж мірою збільшити витривалість, як ПС волокно.
Потреба у кисні. Хоча окиснювальна здатність м'язів визначається кількістю мітохондрій та окиснювальних ферментів у них, окиснювальний метаболізм насамкінець залежить від їх адекватного постачання киснем. У стані спокою потреби організму в АТФ відносно невеликі, тому потреба у кисні також мінімальна. Однак зі збільшенням інтенсивності навантаження зростає і потреба в енергії. Для її задоволення необхідно збільшити окиснювальне утворення АТФ. Задоволення потреб м'язів у кисні здійснюється за рахунок збільшення частоти та глибини дихання, поліпшення процесу газообміну у легенях. Серце починає скорочуватися частіше, постачаючи у м'язи більшу кількість оксигенованої крові.
В організмі людини кисню небагато. Тому кількість кисню, що попадає у кров, котра проходить через легені, прямо пропорційна кількості, використовуваній тканинами для окиснювального фосфорилювання. Отже, можна досить точно визначити величину аеробного продукування енергії, вимірявши кількість кисню, що споживається у легенях.
Дата добавления: 2015-08-26; просмотров: 2045;