Биология культивируемых клеток
1 Рост каллусных клеток, как и целых растений, описывается S-образной кривой. Она получается, если число клеток в культуре или их массу представить как функцию времени, прошедшего с момента их высева (рис. 4). Кривая характерна для популяции клеток, культивируемых как поверхностным способом, так и в жидкой среде, и состоит из следующих фаз: 1)латентная фаза, или лаг-фаза, во время которой отсутствует видимый рост, но идет активный процесс поглощения воды, питательных веществ и подготовки к делению; 2) экспоненциальная, или логарифмическая, фаза роста, в которой по любому принятому критерию возрастает удельная скорость роста; 3) линейная фаза - очень короткая, удельная скорость роста в этой фазе постоянная; 4) фаза замедления роста, в которой скорость роста уменьшается; 5) стационарная фаза, в которой по любому принятому критерию рост постоянен; 6) фаза гибели клетки. Однако форма реальных ростовых кривых может значительно отличаться от моделей продолжительностью фаз. Это зависит как от генотипа (вид растения), так и от условий выращивания и количества инокулюма и транспланта.
Переход клеток из одной фазы в другую контролируется как внутренними, так и внешними факторами. Внутренние факторы: пролиферативный пул, а также продолжительность растяжения, состояние клетки. Внешние факторы: состав питательной среды, уровень рН, содержание кислорода, температура, плотность посева и т.д. Пролиферативный пул – это отношение числа делящихся клеток к общему числа клеток в культуре. Это отношение, выраженное в %, называется митотическим индексом.
|
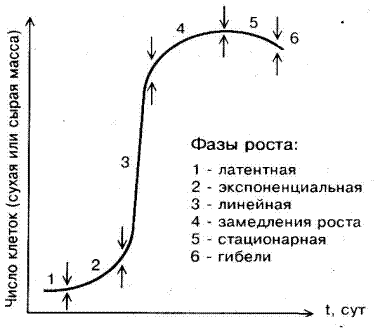
Рисунок 4. Кривая ростового цикла клеток in vitro
В процессе культивирования делятся не все клетки, что приводит к снижению пролиферативного пула. Это может объясняться следующими причинами: 1) необратимая дифференцировка; 2)вступление клеток в G («фаза покоя», или клетки «вне цикла»); 3)гибель клеток. Культура будет находиться в стационарном состоянии в том случае, когда рост клеток делением уравновешивается гибелью части клеток. При этом количество клеток, находящихся в различных фазах, оказывается прямо пропорционально относительной продолжительности данной фазы. Следует отметить, что одиночные клетки в суспензии и их небольшие агрегаты более требовательны к составу питательных веществ, чем каллусные клетки, окруженные себе подобными приповерхностном культивировании. Одиночные клетки в перемешиваемой жидкой среде теряют какие-то вещества, необходимые для деления, поэтому среды для суспензионных культур богаче по своему составу.
2Неорганизованно растущие каллусные клетки, характеризующиеся монотонным размножением и ростом, благодаря процессам вторичной дифференцировки могут образовывать ткани (гистогенез), органы (органогенез), зародышеподобные структуры – эмбриоиды (эмбриоидогенез, или соматический эмбриогенез).
Сначала каллус растет, набирает какую-то определенную критическую массу, потом переходит к морфогенезу. Однако только единичные клетки изменяют свою программу развития. Это говорит о том, что для перехода к морфогенезу необходимо строго определенное сочетание действия внешних факторов. Редифференцировка клетки, связанная с перепрограммированием экспрессии генома – сложный процесс, регулируемый внутренними и внешними факторами. Пока нет ясного представления о природе факторов, благодаря действию которых в массе неорганизованно растущего каллуса возникают организованные структуры - меристематические очаги, дающие начало стеблям, корням, эмбриоидам.
По-видимому, главнейшей причиной, вызывающей дифференцировку клеток и их переход к гистогенезу с последующим формированием органогенных структур, можно считать действие фитогормонов. При этом основным фактором, определяющим тот или иной тип морфогенеза, выступает соотношение ауксинов и цитокининов в питательной среде. Эта закономерность впервые была обнаружена Скугом и Мурасиге, а затем подтверждена многочисленными экспериментами.

Рисунок 5. Регулирование регенерации в культуре тканей сердцевинной паренхимы табака
Как показано на рисунке 5, при среднем соотношении ауксин/цитокинин регенерационных процессов в каллусной ткани табака не наблюдается, а происходит лишь пролиферация клеток. Высокое отношение ауксин/цитокинин вызывает корнеобразование (ризогенез). Низкое отношение ауксин/цитокинин способствует формированию почек (геммогенез). В то же время одна и та же каллусная ткань, находящаяся в пробирке, может дать начало самым различным морфогенным структурам, то есть может иметь место и ризогенез и геммогенез, и формирование соматических эмбриоидов. Эти случаи свидетельствуют о том, что в каллусной ткани возникают концентрационные градиенты фитогормонов, то ест в разных частях каллуса создается различный баланс фитогормонов, индуцирующий тот или иной тип морфогенеза.
Таким образом, тип образующейся меристемы зависит от соотношения между ауксином и цитокинином, но конкретное участие их на разных этапах морфогенеза остается неясным. У целого ряда растений не всегда удается изменением баланса фитогормонов регенерировать почки или корни. На взаимодействие гормонов при регуляции дифференцировки почек или корней могут влиять другие факторы, например содержание сахаров и фосфатов, источник азота и другие компоненты среды, в частности пурины.
На формирование элементов проводящей системы оказывают влияние не только гормоны, но и сахара. При инъекции в каллусную ткань ауксин-сахарного раствора происходит образование сосудистых элементов. Это зависит как от концентрации ауксина, так и от концентрации сахара. Низкие концентрации сахарозы (2 %) вызывали преимущественное образование элементов ксилемы, а высокие (8%) – флоэмы. Использование вместо сахарозы глюкозы не приводили к индукции гистогенеза.
Открытие Скугом и Миллером того факта, что баланс экзогенных фитогормонов регулирует регенерацию почек и корней в тканях каллуса табака, сыграло важную роль в изучении морфогенеза in vitro, но механизм явления на клеточном и молекулярном уровнях остается до сих пор неясным. Отсутствуют фундаментальные знания механизмов индукции морфогенеза под влиянием фитогормонов, поэтому каждый раз при введении в культуру новых растений приходится заново подбирать оптимальный набор и соотношение гормонов.
Для злаков и некоторых бобовых растений не найдены нужные концентрации гормонов для индукции морфогенеза и получения растений-регенерантов из клеток и изолированных протопластов. Подобные факты, а также литературные данные о том, что фитогормоны не обладают специфичностью в регуляции дифференциации и морфогенеза, ставят под сомнение гормональную теорию морфогенеза. Так, Мурасиге отмечает, что концепция Скуга и Миллера слишком упрощает представление об органогенезе. Организованное развитие – сложный процесс, который проходит через ряд этапов, на каждом из них требования к гормонам, свету, температуре заметно отличаются. Кроме того, по мнению Мурасиге, важно учитывать влияние на регенерацию растений веществ, выделяемых самой культурой, которые, являясь токсичными, могут ингибировать морфогенез.
Таким образом, гормональная концепция регуляции дифференциации и морфогенеза in vitro не может рассматриваться как универсальная. Во-первых, фитогормоны в регуляции дифференциации и морфогенеза не обладают специфичностью. Одни и те же гормоны регулируют и деление клеток при недифференцированном росте каллуса, и клеточные деления, связанные с дифференциацией. Во-вторых, есть клетки, не реагирующие на гормональные обработки. Даже если они делятся, тотипотентности не проявляют. В-третьих, способность клеток in vitro проявить свою тотипотентность зависит не только от гормонов. Способность культивируемых тканей к морфогенезу зависит от генотипа исходного растения, его физиологического состояния, органа, из которого изолирован эксплант, времени года и других химических и физических факторов. Существует прямая связь между цитогенетической стабильностью культивируемых клеток и способностью к морфогенезу.
Способы образования эмбриоидов in vitro
Процесс, в котором каллусная клетка имитирует зиготу и дает начало биполярной эмбриоидной структуре, носит название соматического эмбриогенеза, или эмбриоидогенеза. Первыми наблюдали эмбриоидогенез в суспензионной культуре моркови Стюард и Рейнерт в 1958 г. После них это явление было обнаружено у многих видов растений, как в соматических, так и в репродуктивных тканях, однако причины, вызывающие его, пока неясны. Эмбриоидогенез может осуществляться разными путями. Т.Б. Батыгина с коллегами на основе литературных и собственных данных обобщили способы образования эмбриоидов in vitro в виде следующей схемы (рис. 6) . В основе этих способов лежат два пути: образование эмбриоидов непосредственно из одиночной изолированной клетки (1 способ) и из эмбрионального клеточного комплекса (2,3,4 способы). Эмбриональный клеточный комплекс (ЭКК) - это комплекс разнородных клеток, который возникает путем деления одной или нескольких изолированных клеток и в дальнейшем может инициировать эмбриоиды.
 | |||
 | |||
Рисунок 6. Способы образования эмбриоидов in vitro
Возможность возникновения эмбриоида прямо и организованно непосредственно из одиночной изолированной клетки (первый способ) поддерживается не всеми исследователями, так как нет детальных цитоэмбриологических данных по самым начальным этапам эмбриоидогенеза.
Образование эмбриоидов вторым способом показал Рейнер. Он наблюдал как одиночная клетка моркови, выделенная из рыхлого корневого каллуса, помещенная на тонкий слой агара, асимметрично делилась, образуя две дочерние клетки, дальнейшая судьба которых была различна. Одна из них начинала быстро делиться по типу дробления, образовывала шарообразную структуру из мелких прочно связанных клеток - меристематический очаг, то есть эмбриональный клеточный комплекс, который далее дифференцировался в типичный глобулярный или сердечковидный эмбриоид. Вторая дочерняя клетка вытягивалась и образовывала своего рода суспензор. Преобразование всего ЭКК в эмбриоид описали и другие исследователи.
Третий способ образования эмбриоида заключается в формировании его из одной клетки ЭКК, который детально изучал Стрит. Благодаря применению гистохимического, электронно-микроскопического и других методов выявлены особенности строения ЭКК и образования из них эмбриоидов. В суспензии моркови ЭКК состоял из разнородных периферических и центральных клеток. Для поверхностных клеток характерны большое ядро, плотная цитоплазма, мелкие вакуоли, крупные крахмальные зерна. Центральные клетки с толстой стенкой содержат небольшое ядро, крупную вакуоль и много мелких крахмальных зерен. Одна из периферических клеток делится поперечно: апикальная клетка дает начало эмбриоиду, а базальная – суспензору.
Четвертый способ образования эмбриоидов, по мнению Хациус, состоит в формировании его из группы клеток внешнего слоя ЭКК. Однако, из-за методических трудностей не удалось выявить инициальные клетки эмбриоидов. Оказалось возможным определить только слои клеток (поверхностные), дающие начало эмбриоидам. Не обнаружено тонких различий между клетками, вступающими на путь эмбриоидогенеза и не вступающими на него.
В случае культуры клеток и тканей регенерация – это регенерация почек (геммогенез) и корней (ризогенез) или эмбриоидов (эмбриоидогенез). Органогенез и эмбриоидогенез одновременно в одной культуре проявляются крайне редко. Эмбриоиды при переносе на питательную среду без гормонов легко регенерируют целое растение. Увеличив содержание ауксина в среде, можно укоренить почку и тоже получить растение-регенерант. Имеют место случаи одновременной закладки в каллусе апикальных меристем побега и корня (гемморизогенеза), что также приводит к регенерации целого растения. Однако почти невозможно получить растение-регенерант в случае ризогенеза в каллусе (рис. 7).
Следует особо отметить, что для исследований по проблемам морфогенеза и регенерации растений in vitro до сих пор использовались такие удобные модели, как культура тканей табака, моркови, петунии и других видов травянистых двудольных растений. Основные представления о течении и регуляции морфогенеза и регенерации растений in vitro, касаются двудольных, особенно представителей семейства пасленовых.

Рисунок 7. Схема путей морфогенеза in vitro
Явление регенерации доказывает, что дифференциация растительных клеток не означает утрату ими генетических потенций, то есть они остаются тотипотентными. Хотя тотипотентность индивидуальных клеток многих видов растений была продемонстрирована экспериментально, не все исследователи считают, что тотипотентность - свойство, присущее любой растительной клетке. Так, например, трудным объектом в смысле проявления тотипотентности у многих растений оказались клетки зеленых листовых пластинок, одиночные клетки и изолированные протопласты злаков. Каллусообразование и регенерация, вероятно, регулируются различными генетическими механизмами. Генетическая детерминированность регенерационной активности культивируемых клеток является условной, не означающей запрета на возможность регенерировать растения для тех или иных генотипов.
Без регенерации становятся бесполезными все исследования по применению культуры тканей в сельском хозяйстве, то есть биотехнология растений. В настоящее время хорошо разработаны способы регенерации растений in vitro для многих семейств двудольных травянистых растений. Сложнее дело обстоит с травянистыми однодольными растениями, со взрослыми хвойными деревьями.
Таким образом, для понимания механизмов морфогенеза in vitro необходимо их изучение на молекулярном уровне. Генетическая детерминированность морфогенетических процессов означает, что на молекулярном уровне они могут быть представлены как процесс скоординированной в пространстве и времени смены синтезируемых в клетке иРНК и белков.
Дата добавления: 2015-07-14; просмотров: 3966;