Биология культивируемых клеток. 1Переход специализированных неделящихся клеток к пролиферации связан с их дедифференциацией, то есть утратой специализации
1Переход специализированных неделящихся клеток к пролиферации связан с их дедифференциацией, то есть утратой специализации. В основе этого процесса, как и при дифференциации клеток в интактном растении, лежит дифференциальная активность генов. Структура и функции клеток определяются активностью генов, и если клетки различаются по своей структуре и функциям, то это обусловлено различиями в экспрессии их генов, то есть специализация обеспечивается «включением» разных генов в разных клетках. Обычно активна небольшая часть (5 %) всего пула генов, свойственных данному виду. В этот состав активных генов входят, кроме видоспецифичных и обязательных для поддержания клеточного метаболизма, гены, активные только в данном органе, ткани, клетке, а также гены, активные лишь в определенном возрасте или начавшие работать только под влиянием изменившихся внешних условий.
Между геномами в клетках, которые приобретают разную форму и функцию, по-видимому, нет качественных различий, и клетки эти начинают различаться только вследствие разной экспрессии генов. Вновь возникшая клетка обладает широкими потенциями и может развиваться по любому из многих путей в морфологическом и физиологическом смысле.
Детерминация пути развития каждой клетки является основой физиологии развития. Вступление на тот или иной путь развития определяется особым набором белков, то есть каждая специализированная клетка вырабатывает только ей свойственные белки, что является следствием дифференциальной активности генов – экспрессии одной группы генов при одновременной репрессии других.
Белки определяют и фенотип, и функции клетки. Свойства клетки обусловлены действием ферментов и других белков, структура которых определяется и-РНК, образующимися в процессе транскрипции. В то время как структура и метаболизм клетки меняется при дифференциации, набор генов клетки остается неизменным на протяжении всего развития организма. Изменения затрагивают лишь степень активности генов. Из этого следует, что для того, чтобы понять, как происходит дифференциация, нужно выяснить вопрос: почему в клетках одного типа активны одни гены, а в других – другие гены? В процессе развития регуляции белкового синтеза может осуществляться на любом уровне. Выделяют шесть уровней:
1) на уровне синтеза, или репликации ДНК;
2) на уровне синтеза и-РНК, или транскрипции (время и характер образования первичного РНК-транскрипта);
3) на уровне процессинга (характер процессинга первичного РНК-транскрипта);
4) на уровне транспорта и-РНК из ядра в цитоплазму;
5) на уровне трансляции на рибосомах;
6) на уровне деградации и-РНК в цитоплазме.
Регуляция на уровне синтеза ДНК может привести к неравной репликации генов, так что набор генов в разных клетках окажется различным. Активность генов зависит от изменений, происходящих в состоянии ДНК. Установлены три типа устойчивых изменений ДНК, влияющих на активность генов. Во-первых, изменения взаимного расположения генов в хромосомах могут влиять на их функцию. Явление перемещения генов («прыгающие гены») открыла Барбара Мак-Клинток в 1942 г., за что была удостоена Нобелевской премии. Мобильные диспергированные генетические элементы могут перемещаться из одной хромосомы в другую, вызывая сверхмутабельность генов или активацию ранее ингибированных генов. Во-вторых, амплификация (увеличение числа какого-либо гена в геноме) также может привести к изменениям в экспрессии генов. Чаще всего подвержены гены р-РНК. В-третьих, возможны качественные изменения в структуре самого гена (мутация).
Регуляция синтеза белка на уровне транскрипции у прокариот достаточно хорошо объясняет модель Жакобо и Моно. Изучение регуляции активности генов у эукариот очень трудно из-за размера и сложности генома. Возможно, дифференциальная активность генов у эукариот может регулироваться на уровне транскрипции путем маскирования и демаскирования определенных участков генома. Р. Бриттен и Э.Дэвидсон разработали свою гипотезу регуляции генов у высших организмов. Они обратили внимание на то, что ядерная РНК намного богаче уникальными последовательностями, чем и-РНК в цитоплазме, и предположили, что экспрессия генов регулируется на том этапе, когда определяется, какой из потенциальных предшественников и-РНК сохраняется, подвергается процессингу и перейдет в цитоплазму. По терминологии Р.Бриттена и Э.Дэвидсона последовательности ДНК, которые кодируют различные м-РНК, р-РНК и т-РНК, попадающие в цитоплазму, называются генами-продюсерами. Каждый из этих генов регулируется соседней последовательностью ДНК – геном-рецептором. Гены –рецепторы активируются при взаимодействии с РНК определенного типа, называемой активаторной РНК. Активаторная РНК синтезируется на последовательности ДНК, называемой геном-интегратором. Предполагается, что активаторная РНК может действовать на несколько разных генов-рецепторов. Таким образом, один интегратор может вызвать синтез целой группы разных ферментов. Ген-интегратор контролируется соседним сенсорным геном. Он представляет собой последовательность ДНК, с которой взаимодействует внешние агенты, то есть индуцирующие факторы.Гену-продюсеру могут соответствовать различные гены-рецепторы. Это означает, что один ген-продюсер может реагировать на различные индукторы через разные сенсорные и интеграторные гены. Это объясняет факт, как один фермент может быть индуцирован различными стимулирующими веществами. Сложность регуляции синтеза белка также увеличивается за счет того, что одному сенсорному гену может соответствовать несколько генов-интеграторов. Модель регуляции экспрессии генов Р.Бриттена и Э.Дэвидсона хорошо объясняет, как один индуцирующий фактор может вызвать множество различных реакций и в то же время как одинаковые реакции вызываются различными индуцирующими факторами. Трансляция специфических и-РНК начинается не сра- зу после их перехода из ядра в цитоплазму. Важным эволюционным приобретением эукариот являются информосомы, обеспечивающие регуляцию синтеза белка.
Таким образом, дифференциация у растений определяется различиями в экспрессии генов в тех или иных частях растения или на тех или иных стадиях его жизненного цикла. Однако механизмы, контролирующие экспрессию генов по всей цепочке процессов от считывания информации, закодированной в генах до образования активного белка, а также природа и способ воздействия на активацию или регрессию генов, необходимо для дифференцировки и развития, остаются неясными. Способность одной единственной зрелой соматической клетки дать начало целому организму (тотипотентность) показывает, что в процессе нормальной клеточной дифференциации у растений не происходит утраты или необратимой инактивации каких-либо генов. У растений всякая дифференциация обратима при условии, если дифференцированная клетка живая, в протопласте сохранилось ядро и не образовалась вторичная оболочка. Даже такие специализированные клетки, как микроспоры, с помощью ряда экспериментальных процедур можно заставить пролиферировать и дать начало целому организму. Итак, в определенных условиях многие из зрелых растительных клеток сохраняют способность делиться, а в некоторых случаях даже вступить на новый путь развития. Однако вопрос о том, как это происходит, какие события на молекулярном уровне сопровождают этот процесс остается открытым. В образовании каллуса на экспланте принимают участие клетки тканей разных типов, процесс дедифференциации у них, по-видимому, идет по-разному, и возникающие из них каллусные клетки также будут отличаться . Моделью для изучения механизма дедифференциации и каллусообразования лучше всего служит культура сердцевинной паренхимы стебля табака (СПТ), потому что у нее существует строгая зависимость индукции деления и каллусообразования от присутствия ауксина и цитокинина. Обязательным условием дедифференцировки клетки и ее превращения в каллусную клетку является присутствие в питательной среде фитогормонов. Первый этап дедифференциации специализированной клетки – это возобновление способности к делению Р.Г. Бутенко с сотрудниками показали, что дедифференцировка клеток паренхимы в культуре тканей табака начиналась с использования запасных питательных веществ и разрушения специализированных клеточных органелл. Через 6-12 ч после индукции дедифференцировки наблюдались также характерные изменения в тонкой структуре клеток: разрыхление и разбухание клеточной стенки, увеличение числа рибосом, элементов эндоплазматической сети и аппарата Гольджи, увеличение числа и размеров ядрышек. В клетках усиливался синтез транспортных и р-РНК, начинается синтез ДНК, возрастало содержание вновь синтезированных белков. Характерен факт появления специфических белков – антигенов, несвойственных исходной ткани, особенно белка, идентичного одному из белков меристемы растения табака. Все эти изменения происходили только в при
сутствии ауксина и цитокинина и предшествовали делениям, которые начинались через 48-72 часа. Добавление к питательной среде только ауксина, хотя и усиливало синтезы РНК и ДНК, но не вызвало деления клеток, после четырехдневной латентной фазы они переходили к росту растяжением. Присутствие в питательной среде одного кинетина не вызывало изменений в синтезе нуклеиновых кислот. Для осуществления всех процессов, необходимых для деления клеток, требовалось действие кинетина, следующее за ауксином или совместное с ним. Напрашивается вывод, что именно кинетин запускает процесс деления.
Р. Бутенко предполагает следующую цепь событий при этом. Под влиянием стимулятора пролиферации (ауксин) увеличивается проницаемость наружной клеточной мембраны плазмалеммы и изменяются ее свойства. За этим следует активация синтеза цитоплазматических белков на предшествующих ли вновь синтезированных матричных РНК. Эти белки поступают из цитоплазмы в ядро, специфически связываются с гистонами, депрессируя локусы, отвечающие за синтез р- и т- РНК. Репликация ДНК начинается только после достижения некоторого порогового уровня РНК в клетке. Отсутствие кинетина блокирует дальнейшее прохождение клеткой митотического цикла. Очевидно, кинетин определяет синтез специфических РНК и белков, необходимых для перехода клетки к митозу и завершению деления. Существование такого белка –инициатора деления, синтезируемого в премитотическом периоде, и соответствующей ему и-РНК доказано для животных.
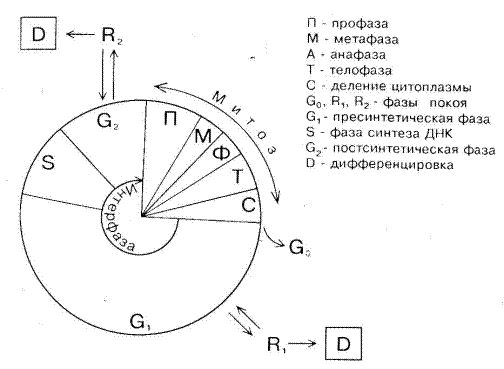
Таким образом, превращение любой клетки в каллусную предшествует процесс глубокой биохимической и структурной перестройки.Однако в деталях тонкие механизмы процесса дедифференцировки растительных клеток не изучены. На рисунке 3 схеме клеточного цикла показано, в какие его фазы клетки могут выйти из цикла деления и перейти в дифференцированное состояние (Д). После митоза и цитокинеза дочерние клетки вступают в интерфазу нового цикла. Интерфаза начинается с фазы G1, во время которой возобновляются интенсивные биосинтетические процессы, резко замедленные во время митоза. Фаза S – это период синтеза ДНК, она заканчивается, когда содержание ДНК в ядре удвоится и хромосомы полностью реплицируются (каждая хромосома состоит из двух идентичных сестринских хроматид). Затем клетки вступают в фазу G2, которая кончается с началом митоза. В начале митоза удвоенные хромосомы, находившиеся ранее в диспергированном интерфазном состоянии, конденсируются и становятся хорошо видимыми в световой микроскоп. Ядерная оболочка разрушается и происходят координированные перемещения хромосом, приводящие к разделению пар сестринских хроматид. После окончания деления ядра образуются две новые ядерные оболочки, а затем делится цитоплазма, в результате чего получаются две дочерние клетки, имеющие по одному ядру. Процесс цитокинеза завершает митоз, начинается интерфаза следующего клеточного цикла. После деления дочерние клетки переходят к фазе растяжения. Растяжение осуществляется благодаря поглощению воды, при этом происходит также новообразование белков, НК, углеводов, жиров и др. веществ. В разделившейся эмбриональной клетке начинается образо-вание компонентов клеточной стенки и цитоплазмы. После растяжения наступает этап перехода клетки к дифференцировке, то есть к узкой специализации.
Клетки переходят к дифференциации (Д) из фазы G1 и G2 , предварительно пройдя в фазу покоя R1и R2. В этих же фазах клеточного цикла они возвращаются в цикл при дифференцировке и индукции их к делению.
Таким образом, после деления перед каждой дочерней клеткой открывается одна из трех возможностей. Клетка может оставаться эмбриональной и вновь вступить в клеточный цикл с последующим митозом либо может оказываться как бы «вне цикла» (Gо), перестав делиться, и наконец, приобретя компетенцию, постепенно детерминироваться и вступить на путь дифференцировки (специализации).
Компетенция – способность клетки воспринимать индуцирующее воздействие и специфически реагировать на него изменением развития. Индуцирующее воздействие могут оказывать различные факторы: гормоны, продукты жизнедеятельности соседних клеток, других тканей, электрофизиологические сигналы и т.д.
Детерминация – приобретение клеткой состояния готовности к реализации определенных наследственных свойств. Детерминация приводит к развитию по определенному пути с одновременным ограничением возможности развития в других направлениях. Детерминация компетентной клетки может начинаться сразу же после деления в начале роста протоплазмы. Детерминированная определенным образом клетка приобретает узкую специализацию, то есть дифференцируется и превращается в клетку какой-либо ткани.
Получить каллус первый раз из нового растительного материала бывает трудно, потому что ткани разных видов растений и даже разных частей растения требует для дедиффренциации и каллусогенеза разных питательных сред, особенно это касается количества и соотношения гормонов. При длительном пассировании ткани некоторых растений приобретают способность к синтезу достаточно больших количеств ауксина, а также цитокинина.
2 Основным типом культивируемых клеток являются каллусные. В цикле выращивания каллусные клетки после ряда делений проходят обычный для клетки растения онтогенез, т.е. приступают к росту растяжением, затем дифференцируются как зрелые каллусные клетки, стареют и гибнут. Дифференцированные каллусные клетки специализируются на синтезе видоспецифических вторичных соединений. Эти клетки подобны паренхимных клеткам растения. В растущем каллусе делению подвергаются не все клетки, это функция мерис-теподобных клеток с густой плотной цитоплазмой и без вакуолей.
| |||
|
Любой фрагмент растения представляет собой мозаику различных тканей, и в зависимости от того, какая ткань даст начало каллусу, возникшие даже из одинаковых эксплантов каллусы будут гетерогенными отличающимися друг от друга. Одинаковых, в полном смысле, эксплантов в природе быть не может, следовательно, неоднородность исходного материала (видовая, возрастная, физиологическая) предопределяет разнокачественность клеток в культуре.
Физиологическая гетерогенность состоит в том, что клетки в популяции находятся в разном физиологическом состоянии, т.е. делятся, растут, стареют, погибают. Такая культура называется асинхронной. Заставить популяцию клеток высших растений проходить фазы клеточного цикла одновременно, т.е. синхронизировать их почти невозможно. Потому что, та часть клеток, которая способна в данный момент к делению, составляет 2-4 %. Неблагоприятные условия (низкая температура, исключение важных компонентов питания), задерживающие деление, в какой-то степени способствуют накоплению числа клеток, готовых к делению. Более эффективны некоторые химические вещества, блокирующие определенные стадии подготовки к делению. В лучших случаях синхронизация может быть достигнута у 10-30 % клеток, но при последующих делениях популяция опять быстро утрачивает синхронность.
Следует подчеркнуть, что физиологическая вариабельность клеток в суспензионной культуре меньше по сравнению с культурой каллусной ткани на агаре, что связано с более однородными условиями питания, аэрации и удаления токсических метаболитов из клеточного окружения в жидкой перемешиваемой среде.
Гетерогенность культивируемых клеток обусловлена генетической, эпигенетической и модификационной изменчивостью. Генетические, или мутационные, изменения приводят к изменению генотипа, которое может быть унаследовано. Мутации (изменения количества или структуры ДНК) происходят на генном, хромосомном и геномном уровнях.
Генная или точечная мутация означает изменение структуры ДНК в одном локусе. Генные мутации приводят к сильным или слабым изменениям морфологических, биохимических и физиологических свойств клетки. Мутации, возникающие в результате изменения макроструктуры хромосом, называются хромосомными мутациями, или хромосомными перестройками (аберрациями). Структурные перестройки хромосом возникают в результате инверсии, делеции, дупликации, транслокации. Геномные мутации связаны с изменением числа хромосом в ядре, т.е с изменениями в кариотипе.
Все виды названных генетических изменений имеют место у клеток in vitro. Наиболее подробно исследована хромосомная изменчивость клеток.
Процесс возрастающего доминирования в популяции клеток определенного типа называется клеточной селекцией. Доминирование может быть вызвано преимущественной пролиферацией одних клеток или успешной элиминацией (удалением) других. Такая селекция называется автоселекцией.
Разнообразие (вариабельность) среди клеточных линий или растений-регенерантов называют сомаклональной вариабельностью.
Клетки in vitro становятся разнокачественными также благодаря эпигенетическим изменениям, то есть изменениям в программе считки генов. Эти изменения генной активности являются наследуемыми.
К ненаследуемым изменениям у клеток в культуре относятся модификационные изменения, которые в большинстве носят адаптивный приспособительный характер. Такие изменения не затрагивают генетических структур клетки, они соответствуют физиологической адаптации, при которой границы изменений не превышают норму реакции, обусловленной генотипом.
Гетерогенность клеток возрастает с увеличением продолжительности их культивирования.
Дата добавления: 2015-07-14; просмотров: 2680;