Ионная теория электрогенеза Бернштейна
Современная теория электрогенеза вытекает исторически из положений Бернштейна (1940), связавшему впервые ионную проницаемость мембран с природой биопотенциалов.
Ее основные положения:
1. В состоянии покоя мембрана проницаема только для ионов калия.
2. Ионы калия, выходя из клетки, создают потенциал, величина которого рассчитывается по уравнению Нернста:
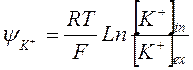
где [К+]i и [К+]e – концентрации внутриклеточных – i и внеклеточных ионов – e, соответственно.
3. Этот потенциал равновесный, так как различие концентраций [К+]i и [К+]e присутствует в клетках постоянно.
В качестве доказательства своей теории Бернштейн указывал на зависимость мембранного потенциала от температуры, изменений внеклеточной концентрации ионов калия [К+]e, т.е. от расчетных показателей уравнения Нернста.
Теория постоянного поля и потенциал покоя (ПП)
Современные представления об электрогенезе основаны на положениях теории постоянного поля. Ее основные создатели: Ходжкин, Хаксли и Катц (1949) разработали ряд упрощений для возможного анализа протекания электрических процессов на мембране:
1. Ионы движутся в мембране по градиенту концентрации и электрического поля, как в растворе.
2. Напряженность электрического поля в мембране постоянна и потенциал падает линейно ее толщины по мере удаления от источника электрического поля.
3. Мембрана гомогенна, имея постоянную диэлектрическую проницаемость по всей толщине.
4. Концентрация ионов в мембране на границе с раствором пропорциональна концентрации ионов в самом растворе.
Из первого выражения вытекает, что плотность ионного тока через мембрану:
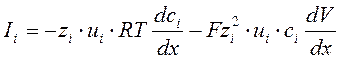 , где:
, где:
zi –валентность i-го иона, V- потенциал в точке, x – расстояние по толщине мембраны от наружней ее границы. Т.е.плотность ионного тока определяется разностью между диффузионным потоком i-го иона по концентрационному градиенту (1-ое слагаемое) и противоположно направленным потоком этого иона, обусловленным электрическим полем, возникшем при его диффузии (2-ое слагаемое).
При постоянстве электрического поля dV/dx =const=V/a, где а – тощина мебраны константа проницаемости мембраны (Pi) определяется относительной подвижностью и растворимостью в мембране:
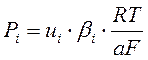 , где:
, где:
βi – коэффициент распределения i-го иона между водным раствором и мембраной, ui –подвижность этого иона в электрическом поле.
На основании представленных положений теории постоянного поля Ходжкин, Гольдман и Катц вывели уравнение зависимости плотности ионных токов от мембранного потенциала и ионной проницаемости:
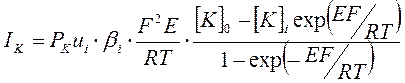
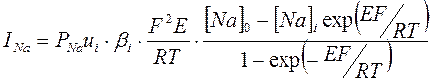
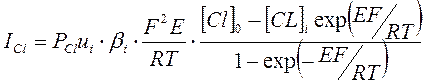
Предполагая, что при потенциале покоя (ПП) суммарный ионный ток равен нулю Ходжкин, Гольдман и Катц вывели уравнение:
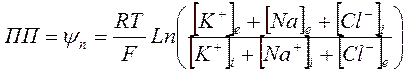 , где:
, где:
[К+]i, [Na+]i, [Сl-]i и [К+]e [Na+]e и [Cl-]e – концентрации внутриклеточных – intra и внеклеточных ионов – extra соответственно,
PK, PNa и PCl – проницаемость соответствующих ионов.
Для ПП гигантского аксона кальмара соотношение:
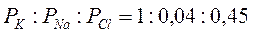
Таким образом, ПП имеет в основном калиевую природу, что и определяет его вклад в изменения мембранного потенциала при модуляции концентрации ионов калия. Распределение ионов между клеткой и средой при ПП устанавливается на постоянном уровне, и суммарный ионный ток равняется нулю:
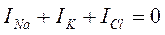
Постоянно существующий градиент, поддерживающий пассивный поток ионов через клеточную мембрану, требует процессов, его восстанавливающих. Их выполняет система активного транспорта – Mg2+-зависимая Na+/K+-АТФ-аза, которая осуществляет противоградиентный транспорт ионов Na+ и K+. Энергозависимый процесс происходит в режиме одновременного выброса 3-х ионов Na+ из клетки и закачивания 2-х ионов K+ внутрь. Создание дополнительного дефицита внутриклеточного содержимого в положительном заряде позволило режим работы данной системы, кроме энергозависимого, считать еще и электрогенным.
Потенциал действия (ПД)
Раздражение, как уже упоминалось, лишь при достижении порога возбуждения вызывает самоподдерживающийся процесс изменения значений биопотенциалов в клетке – потенциал действия. Пороговые условия начала развития ПД определяются многочисленными условиями (законами), подробно рассмотренными в соответствующих электрофизиологических литературных разделах. Например, они включают в себя силу (амплитуду) и длительность раздражающего стимула. Взаимосвязь этих параметров представляет собой известную гиперболическую зависимость "сила – длительность" (Рис.17).
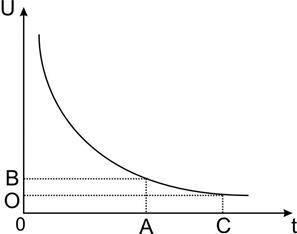
| Рис.17. Зависимость сила – длительность. По оси абсцисс- длительность стимула; по оси ординат -амплитуда стимула С - полезное время; А -хроноксия; О - реобаза |
Такие характеристики, как полезное время, реобаза, хронаксия являются общими для электровозбудимых тканей (там же).
Электротонический потенциал и локальный ответ. Изменения, проходящие на мембране при действии раздражения до порогового уровня, проявляются в виде электротонического потенциала и локального ответа.
Электротонический потенциал представляет собой изменения пассивных (омических и емкостных) электрических характеристик мембран в ответ на раздражение, амплитуда которого меньше 50 – 70% пороговых (100%) значений.
Локальный ответ представляет собой изменения подпороговых активных (ионная проницаемость) электрических характеристик мембран в ответ на раздражение, амплитуда которого ниже пороговых значений.
Таким образом, основной причиной инициации процесса развития ПД является достижение критического (порогового) уровня смещения мембранного потенциала.
Дата добавления: 2015-06-22; просмотров: 1794;