Ионная природа потенциала действия (ПД). Формальное описание ионных токов
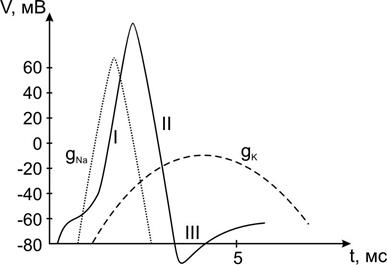
ПД, регистрируемый впервые на гигантском (до 500 мкм в диаметре) аксоне кальмара, состоит из нескольких фаз (Рис. 11).
|
Исходно от уровня потенциала покоя (-90 мВ) начинается I-я фаза деполяризации, сменяющаяся на уровне нулевого (0 мВ) мембранного потенциала противоположным знаком овершутом (+ 40 мВ) и затем переходящая в II-ю фазу реполяризации по пути возвращения значений мембранного потенциала к потенциалу покоя. Отклонения от пути возвращения называют III-й фазой – следовым потенциалом:
А) положительным – при продолжающейся реполяризации.
Б) отрицательный – при развитии деполяризации.
Метод фиксации потенциала и модификации ионного состава растворов позволили вскрыть ионные механизмы каждой фазы ПД.
Основное участие в развитии фазы деполяризации принимает входящий в клетку поток положительных ионов натрия (Na+), перезаряжающих внутреннюю поверхность мембраны. На смену быстрой активации натриевой проницаемости пороговым раздражителем приходят процессы инактивации входа Na+ и активации выхода из клетки ионов калия (K+), что проявляется фазой реполяризации – возвращения зарядов на внутренней поверхности мембраны к отрицательным значениям.
С помощью ряда упрощений Ходжкину и Хаксли (1950) в виде уравнений удалось произвести формальное (математическое) описание кинетики ионных токов электровозбудимой мембраны. По их мнению ионный ток (I) складывается из суммы натриевого (INa) калиевого (IK) и тока утечки (Il):
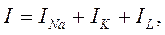
В отличие от натриевого и калиевого тока, ток утечки не подчиняется потенциал-зависимым механизмам активации и инактивации.
Каждый из токов рассчитывается по закону Ома:
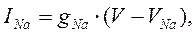
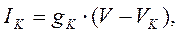
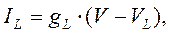
где: gNa, gK и gl – проводимость для ионов натрия, калия и ионов утечки соответственно. (V-V(Na,K,L)) – величины электрохимических потенциалов для соответствующих ионов, где V – является отклонением от абсолютных значений мембранного потенциала Е, а V(Na,K,L)) – равновесные потенциалы, рассчитанные по уравнению Нернста.
С помощью выражений:
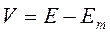 и соответственно:
и соответственно: 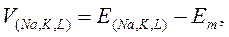 откуда:
откуда:
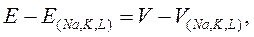
Для гигантского аксона кальмара:
ЕNa=+55мВ, ЕK= -72мВ, ЕL= -50мВ.
В свою очередь, величины ионных проводимостей мембраны равны:
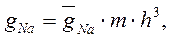
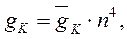
где: gNa и gK – максимальные проводимости мембраны (при сильной деполяризации ĝ Na = 120 ммо/см2, ĝК = 36 ммо/см2, gl = 0,3 ммо/см2) для ионов натрия и калия соответственно.Величины m и n - переменные процесса активации, h - переменная инактивации.
Их значения в зависимости от мембранного потенциала изменяются в пределах от 0 до 1 и рассчитываются из системы дифференциальных уравнений:
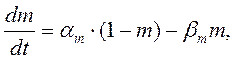
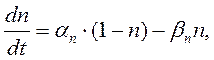
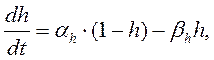
где am, bm, an, bn, ah, bh- константы скоростей, зависящие от мембранного потенциала, температуры и концентрации двухвалентных ионов в наружном растворе, но не от времени. При деполяризации мембраны значения am, an и bh увеличиваются, а bm, bn и ah- уменьшаются. Решения этих уравнений проще представить в виде экспоненциальных характеристик – постоянных времени изменения m, n и h:
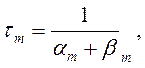
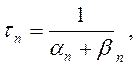
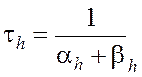
Стационарные значения переменных m, n и h будут равны:
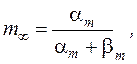
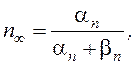
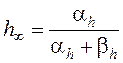
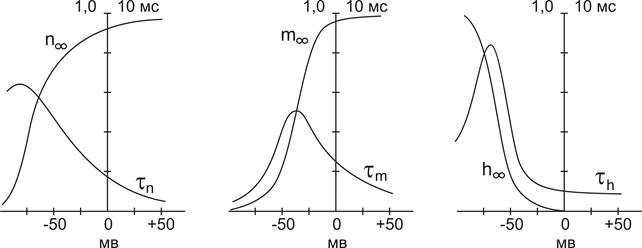
Графики зависимости стационарных значений m, n и h (m¥, n¥ и h¥) и постоянных времени tm,tn и th от мембранного потенциала представлены на Рис. 12
|
На основании представленных выше теоретических выкладок Ходжкиным и Хаксли были рассчитаны параметры потенциала действия, которые сравнивались с экспериментальными.
В состав мембранного тока (Im) кроме ионной компоненты входит и емкостная составляющая: 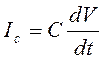 и тогда:
и тогда:
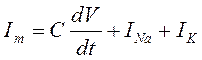
При поддержании потенциала на постоянном уровне (метод clamp-voltage) емкостная составляющая исчезает, и мембранный ток удается зарегистрировать как сумму натриевого и калиевого ионных токов (Рис.)
Использование блокаторов ионных каналов позволило получить отдельные вольт-амперные характеристики для натриевого и калиевого ионных токов и вскрыть ионные механизмы развития потенциала действия. На Рис. видно, что отличия рассчитанных и экспериментальных параметров незначительны.
Показанные изменения ионных проницаемостей (а) и ионных токов (б), рассчитанные с помощью уравнений Ходжкина-Хаксли при возникновении ПД в гигантском аксоне кальмара (Рис.) в ответ на очень короткий стимул позволили избежать емкостной составляющей мембранного тока. Видно, что первый нуль Ii соответствует моменту, когда входящий натриевый ток становится равным выходящему ионному току Il+IK. Это момент критической деполяризации, когда локальный ответ начинает переходить в ПД. В начале этого периода PNa и INa совпадают с началом развития ПД, однако, затем ход изменений их становится разным:
1. Изменения PNa имеют одну вершину, совпадающую в максимальной точке с вершиной ПД.
2. Кривая изменений INa характеризуется двумя максимумами, из которых один приходится на примерно на середину восходящего ПД, а второй- на первую треть фазы реполяризации. Вершине ПД соответствует точка наибольшего падения кривой INa в области “седла” между двумя ее вершинами.
Различия в динамике изменения между PNa и INa обусловлены тем, PNa является прямым следствием деполяризации в момент ПД, а INa зависит также от электрохимического потенциала (V-VNa), величина которого по мере деполяризации снижается. В результате продолжающего компенсаторного возрастания PNa, INa продолжает нарастать до момента выравнивания V и VNa, когда суммарный ионный ток (Ii) и изменения ПД достигают максимума. Наличие емкостных свойств мембраны позволяет ПД нарастать еще некоторое время, даже в отсутствии усиления Ii. Остановка в нарастании ПД будет происходить при равенстве плотностей выходящих и входящих ионных токов, что соответствует максимальной величине ПД.
Преобладание выходящей компоненты ионных токов будет приводить развитию фазы реполяризации, которая будет замедляться вторичным повышением INa, связанным с увеличением электрохимического потенциала (V-VNa) при еще достаточно высоком PNa. В фазу реполяризации PNa падает сначала круто, а затем, более полого из-за усиления инактивации деполяризованной мембраны.
Расчеты, проведенные позже на миелиновых нервных волокнах в области перехвата Ранвье показали, что кинетика изменений ионных проницаемостей в момент развития ПД качественно не отличается от таковой в гигантском аксоне кальмара (см.Выше). Следует отметить характерную для перехватов Раньве более высокую (3 раза) скорость нарастания ПД. Известные же отличия в скорости проведения по мякотным и безмякотным нервным волокнам определяются, как показывают дальнейшие расчеты, параметрами сопротивления и емкости мембраны с таковыми в перехватах Ранвье. При высоких сопротивлениях миелиновой оболочки (до ГОм·см2), в области перехватов низко сопротивление мембраны (см. Ниже).
Дата добавления: 2015-06-22; просмотров: 4118;